Таблица эволюция головного мозга у животных. Этапы эволюции головного мозга позвоночных
Государственное образовательное учреждение высшего профессионального образования «Ставропольская государственная медицинская академия» Министерства здравоохранения и социального развития Российской ФедерацииКафедра биологии с экологией
А.Б.Ходжаян
Н.Н. Федоренко
Л.А. Краснова
К НЕКОТОРЫМ ВОПРОСАМ ЭВОЛЮЦИИ
(дополненное)
Методическое пособие для студентов 1-го курса СтГМА
СТАВРОПОЛЬ,
УДК 57:575.
К некоторым вопросам эволюции. Методическое пособие для студентов 1 курсов. Изд: СтГМА. 2009г.с.31.
В учебнике биологии под ред. А.А.Слюсарева и С.В. Жуковой, которым пользуются студенты 1-го курса при изучении медицинской биологии и генетики, некоторые вопросы теории эволюции требуют дополнения и уточнения. Сотрудники кафедры биологии СтГМА сочли необходимым составить данное методическое пособие по некоторым вопросам теории эволюции живой природы.
Составители: д.м.н., проф. Ходжаян А.Б.,
К.м.н., доц. Федоренко Н.Н.,
К.м.н., доц. Краснова Л.А.
© Ставропольская государственная
медицинская академия, 2009
^ ФИЛОГЕНЕЗ СИСТЕМ ОРГАНОВ У ЖИВОТНЫХ
Основы строения и функции различных органов и систем органов у животных и человека не могут быть достаточно глубоко и полно поняты без знания их исторического становления, то есть филогенеза.
^ Филогенез нервной системы .
Все живые организмы на протяжении жизни испытывают многообразные воздействия со стороны внешней среды, на которые отвечают изменением поведения или физиологических функций. Эта способность реагировать на средовые воздействия называется раздражимостью.
Раздражимость имеет место уже у простейших и выражается в изменении у них процессов жизнедеятельности или поведения в ответ на такие раздражения, как химические, температурные, световые.
У многоклеточных животных появляется специальная система клеток - нейроны, способные в ответ на определенные раздражения отвечать нервным импульсом, который они передают другим клеткам тела. Совокупность нервных клеток формирует нервную систему, сложность структуры и функции которой возрастает с усложнением организации животных. В зависимости от последнего у многоклеточных животных в эволюции сложилось три основных типа нервной системы: сетевидная (диффузная), ганглиозная (узловая) и трубчатая.
^
Д иффузная (сетевидная
)
нервная
система характерна для наиболее примитивных животных - кишечнополостных. У них нервная система состоит из диффузно расположенных по всему телу нейронов, которые своими отростками контактируют друг с другом и с иннервируемыми ими клетками, образуя подобие сети. Такой тип организации нервной системы обеспечивает высокую взаимозаменяемость нейронов и тем самым большую надежность функционирования. Однако ответные реакции при таком типе организации нервной, системы имеют неточный, расплывчатый характер.
иффузная (сетевидная
)
нервная
система характерна для наиболее примитивных животных - кишечнополостных. У них нервная система состоит из диффузно расположенных по всему телу нейронов, которые своими отростками контактируют друг с другом и с иннервируемыми ими клетками, образуя подобие сети. Такой тип организации нервной системы обеспечивает высокую взаимозаменяемость нейронов и тем самым большую надежность функционирования. Однако ответные реакции при таком типе организации нервной, системы имеют неточный, расплывчатый характер.
^ Узловой (ганглиозный) тип является следующей ступенью в развитии нервной системы. Он характерен для всех червей, иглокожих, моллюсков и членистоногих. У них имеет место концентрация тел нейронов в виде единых скоплений - узлов (ганглиев). Причем, у плоских и круглых червей такие узлы располагаются только на переднем конце тела, где сосредоточены органы захвата пищи и органы чувств. У кольчатых червей и членистоногих, тело которых разделяется на сегменты, кроме головных узлов, формируется брюшная цепочка нервных узлов, которые регулируют работу тканей и органов данного сегмента (кольчатые черви) или группы сегментов (членистоногие). Однако, всегда наиболее развитым остается головной узел, являющийся координирующим и регулирующим центром по отношению к остальным ганглиям. Этот тип нервной системы характеризуется некоторой организованностью: где возбуждение проходит строго по определенному пути, что дает выигрыш в быстроте и точности реакции. Но этот тип нервной системы весьма раним.
Хордовым животным присущ трубчатый тип нервной системы. У них в эмбриональном периоде из эктодермы над хордой закладывается нервная трубка, которая у ланцетника сохраняется всю жизнь и выполняет функцию центрального отдела нервной системы, а у позвоночных она преобразуется в спинной и головной мозг. При этом головной мозг развивается из переднего отдела нервной трубки, а из остальной ее части - спинной мозг.
Головной мозг у позвоночных состоит из пяти отделов: переднего, промежуточного среднего, продолговатого мозга и мозжечка.

^ ЭВОЛЮЦИЯ ГОЛОВНОГО МОЗГА У ПОЗВОНОЧНЫХ
Формирование головного мозга у зародышей всех позвоночных начинается с появления на переднем конце нервной трубки вздутий - мозговых пузырей. Вначале их образуется три, а затем пять. Из переднего мозгового пузыря в дальнейшем образуется передний и промежуточный мозг, из среднего - средний мозг, а из заднего - мозжечок и продолговатый мозг. Последний без резкой границы переходит в спинной мозг
В нервной трубке есть полость - невроцель, которая в ходе образования пяти мозговых пузырей формирует расширения - мозговые желудочки (у человека их 4).В этих участках мозга различают дно (основание) и крышу (мантия). Крыша располагается над -, а дно под желудочками.
Вещество мозга неоднородно - представлено серым и белым веществом. Серое - это скопление нейронов, а белое образовано отростками нейронов, покрытыми жироподобным веществом (миелиновой оболочкой), которое придает веществу мозга белый цвет. Слой серого вещества на поверхности крыши любого отдела мозга называется корой.
Большую роль в эволюции нервной системы играют органы чувств. Именно концентрация органов чувств на переднем конце тела обусловила прогрессивное развитие головного отдела нервной трубки. Полагают, что передний мозговой пузырь сформировался под влиянием обонятельного, средний - зрительного, а задний - слухового рецепторов.
^ Рыбы
Передний мозг небольшой, не разделен на полушарии, имеет только один желудочек. Его крыша не содержит нервных элементов, а образована эпителием. Нейроны сосредоточены на дне желудочка в полосатых телах и в отходящих спереди от переднего мозга обонятельных долях. По существу, передний мозг выполняет функцию обонятельного центра.
^ Средний мозг является высшим регуляторным и интегративным центром. Он состоит из двух зрительных долей и является наиболее крупным отделом мозга. Такой тип мозга, где высшим регуляторным центром является средний мозг, называется ихтиопсидпым .
^ Промежуточный мозг состоит из крыши (таламуса) и дна (гипоталамуса) С гипоталамусом связан гипофиз, а с таламусом - эпифиз.
Мозжечок у рыб хорошо развит, поскольку их движения отличаются большим разнообразием.
^ Продолговатый мозг без резкой границы переходит в спинной мозг и в нем сосредоточен пищевой, сосудодвигательный и дыхательный центры.
От мозга отходит 10 пар черепно-мозговых нервов, что характерно для низших позвоночных.
Амфибии
У амфибий имеется ряд прогрессивных изменений в головном мозге, что связано с переходом к наземному образу жизни, где условия по сравнению с водной средой более разнообразны и характеризуются непостоянством действующих факторов. Это привело к прогрессивному развитию органов чувств и соответственно - прогрессивному развитию головного мозга.
^ Передний мозг у амфибии в сравнении с рыбами значительно крупнее, в нем появилось два полушария и два желудочка. В крыше переднего мозга появились нервные волокна, образующие первичный мозговой свод - архипаллиум . Тела нейронов располагаются в глубине, окружая желудочки, в основном в полосатых телах. Все еще хорошо развиты обонятельные доли.
Высшим интегративным центром остается средний мозг (ихтиопсидный тип). Строение такое же, как у рыб.
Мозжечок связи с примитивностью движений амфибий имеет вид небольшой пластинки.
^ Промежуточный и продолговатый мозг такие же, как у рыб. От головного мозга отходят 10 пар черепно-мозговых нервов.
Пресмыкающиеся (рептилии)
Рептилии относятся к высшим позвоночным и характеризуются более активным образом жизни, что сочетается с прогрессивным развитием всех отделов головного мозга.
^ Передний мозг является самым крупным отделом головного мозга. Кпереди от него отходят развитые обонятельные доли. Крыша остается тонкой, но на медиальной и латеральной стороне каждого полушария появляются островки коры. Кора имеет примитивное строение и называется древней – археокортекс. Роль высшего интегративного центра выполняют полосатые тела переднего мозга - зауропсидный тип мозга . Полосатые тела обеспечивают анализ поступающей информации и выработку ответных реакций.
^ Промежуточный, мозг, будучи связан с эпифизом и гипофизом имеет еще и дорсальный придаток - теменной орган, воспринимающий световые раздражения.
Средний мозг теряет значение высшего интегративного центра, снижается и его значение как зрительного центра, в связи, с чем его размеры уменьшаются.
Мозжечок развит значительно лучше, чем у амфибий.
^ Продолговатый мозг образует резкий изгиб, характерный для высших позвоночных, в том числе и для человека.
От головного мозга отходит 12 пар черепно-мозговых нервов, что типично для всех высших позвоночных, в том числе и для человека.
Птицы
Нервная система в связи с общим усложнением организации, приспособленностью к полету и обитанием в самых различных средах, развита значительно лучше, чем у пресмыкающихся.
Дня птиц характерно дальнейшее увеличение общего объема головного мозга, особенно переднего.
^ Передний мозг у птиц - это высший интегративный центр. Его ведущим отделом являются полосатые тела (зауропсидный тип мозга).
Крыша остается слабо развитой. В ней сохраняются только медиальные островки коры, которые выполняют функцию высшего обонятельного центра. Они оттесняются к перемычке между полушариями и носят название гиппокампа. Обонятельные доли развиты слабо.
^ Промежуточный мозг небольших размеров и связан с гипофизом и эпифизом.
Средний мозг имеет хорошо развитые зрительные доли, что обусловлено ведущей ролью зрения в жизни птиц.
Мозжечок крупный, имеет среднюю часть с поперечными бороздами и небольшие боковые выросты.
^ Продолговатый мот такой же, как у рептилий. 12 пар черепно-мозговых нервов.
Млекопитающие
Передний мозг - это наиболее крупный отдел головного мозга. У разных видов его абсолютный и относительный размеры весьма варьируют. Главная особенность переднего мозга - значительное развитие коры полушарий, которая собирает всю сенсорную информацию от органов чувств, производит высший анализ и синтез этой информации и становится аппаратом тонкой условно-рефлекторной деятельности, а у высокоорганизованных млекопитающих - и психической деятельности (маммальныйтип мозга).
У наиболее высокоорганизованных млекопитающих кора имеет борозды и извилины, что значительно увеличивает ее поверхность.
Д ля переднего мозга млекопитающих и человека характерна функциональная асимметрия. У человека, она выражается в том, что правое полушарие отвечает за образное мышление, и левое - за абстрактное. Кроме того, в левом полушарии находятся центры устной и письменной речи.
ля переднего мозга млекопитающих и человека характерна функциональная асимметрия. У человека, она выражается в том, что правое полушарие отвечает за образное мышление, и левое - за абстрактное. Кроме того, в левом полушарии находятся центры устной и письменной речи.
^ Промежуточный мозг содержит около 40 ядер. Специальные ядра таламуса перерабатывают зрительные, тактильные, вкусовые и интероцептивные сигналы, направляя их затем в соответствующие зоны коры больших полушарий.
В гипоталамусе сосредоточены высшие вегетативные центры, управляющие работой внутренних органов через нервные и гуморальные механизмы.
В среднем мозге на смену двухолмия приходит четверохолмие. Его передние холмы являются зрительными, а задние связаны со слуховыми рефлексами. В центре среднего мозга проходит ретикулярная формация, которая служит источником восходящих влияний, активирующих кору больших полушарий. Хотя передние доли являются зрительными, анализ зрительной информации осуществляется в зрительных зонах коры, а на долю среднего мозга приходится главным образом управление глазной мускулатурой - изменение просвета зрачка, движения глаз, напряжение аккомодации. В задних холмах расположены центры, регулирующие движения ушных раковин, натяжение барабанной перепонки, перемещение слуховых косточек. Средний мозг также участвует в регуляции тонуса скелетной мускулатуры.
Мозжечок имеет развитые боковые доли (полушария), покрытые корой, и червь. Мозжечок связан со всеми отделами нервной системы, имеющими отношение к управлению движениями - с передним мозгом, стволом мозга и вестибулярным аппаратом. Он обеспечивает координацию движений.
^ Продолговатый мозг . В нем по бокам обособляются пучки нервных волокон, идущих к мозжечку, а на нижней поверхности - продолговатые валики, получившие название пирамид.
От основания мозга отходит 12 пар черепно-мозговых нервов.
^ ФИЛОГЕНЕЗ КРОВЕНОСНОЙ СИСТЕМЫ
У многоклеточных организмов клетки теряют непосредственный контакт с окружающей средой, в связи, с чем возникает необходимость в появлении системы транспорта жидкостей для доставки к клеткам необходимых веществ и удаления продуктов жизнедеятельности. У низших беспозвоночных (губок, кишечнополостных, плоских и круглых червей) транспорт веществ происходит путем диффузии токов тканевой жидкости. У более высокоорганизованных беспозвоночных, а также у хордовых, появляются обеспечивающие циркуляцию веществ сосуды. Возникает кровеносная, затем лимфатическая система. И та, и другая развиваются из мезодермы.
Эволюционно сложилось два типа кровеносной системы: замкнутая и незамкнутая . В замкнутой кровь циркулирует только по сосудам, а в незамкнутой часть пути она проходит по щелевидным пространствам - лакунам и синусам.
Впервые кровеносная система появляется у кольчатых червей. Она замкнутая. Сердца еще нет. Имеются два главных продольных сосуда - брюшной и спинной, связанные между собой несколькими кольцевыми сосудами, идущими вокруг кишечника. От главных сосудов отходят более мелкие сосуды к органам, движение крови идет по спинному сосуду вперед, а по брюшному - назад.
У членистоногих кровеносная система достигает более высокой организации. У них имеется центральный пульсирующий аппарат - сердце, оно расположено на спинной стороне тела, При его сокращении кровь поступает в артерии, откуда изливается в щелевидные пространства между органами (синусы и лакуны), а затем вновь всасывается через парные отверстия в сердце то кровеносная система у членистоногих незамкнутая.
У насекомых кровь не выполняет функцию транспорта газов, обычно она бесцветна и называется гемолимфой.
У моллюсков кровеносная система тоже незамкнутая, но у них, кроме артерий, есть и венозные сосуды. Сердце имеет несколько предсердий, куда впадают вены, и один крупный желудочек, от которого отходят артерии.
У наиболее примитивных хордовых животных - у ланцетника, кровеносная система во многом напоминает сосудистую систему кольчатых червей, что говорит об их филогенетическом родстве. У ланцетника нет сердца, его функцию выполняет брюшная аорта. По ней течет венозная кровь, которая поступает в жаберные сосуды, обогащается кислородом, а затем идет в спинную аорту, несущую кровь ко всем органам. Венозная кровь из передней части тела собирается в передние, а из задней - в задние кардинальные вены. Эти вены сливаются в кювьеровы протоки, по которым кровь поступает в брюшную аорту.
В эволюции позвоночных наблюдается появление сердца, расположенного на грудной стороне тела, и усложнение его строения от двухкамерного до четырехкамерного. Так у рыб сердце состоит из одного предсердия и одного желудочка, в нем течет венозная кровь. Круг кровообращения один и кровь не смешивается. Круговорот крови во многом сходен с кровеносной системой ланцетника.
У наземных позвоночных в связи с приобретением легочного дыхания развивается второй круг кровообращения и сердце, кроме венозной, начинает получать артериальную кровь. При этом система сосудов дифференцируется на кровеносную и лимфатическую.
Промежуточную ступень в развитии системы кровообращения от низших позвоночных к высшим занимает кровеносная система земноводных и пресмыкающихся. У этих животных имеется два круга кровообращения, но в сердце происходит смешивание артериальной и венозной крови.
Полное разделение артериальной и венозной крови характерно для птиц и млекопитающих, у которых четырехкамерное сердце. Из двух дуг аорты, характерных для амфибий и рептилий, остается только одна: у птиц - правая, а у млекопитающих - левая.
^ Эволюция артериальных дуг.
У эмбрионов всех позвоночных животных впереди от сердца закладывается непарная брюшная аорта, от которой отходят артериальные дуги. Они гомологичным артериальным дугам ланцетника. Но число их меньше, чем у ланцетника: у рыб - 6-7 пар, а у наземных позвоночных 6 пар.
Первые две пары у всех позвоночных испытывают редукцию. Следующие пары артериальных дуг у рыб разделяются на приносящие и выносящие жаберные артерии, а у наземных животных подвергаются сильным преобразованиям. Так, из 3-й пары дуг образуются сонные артерии. Четвертая пара преобразуется в дуги аорты, которые у амфибий и рептилии развиваются симметрично. У птиц левая дуга атрофируется и сохраняется только правая. У млекопитающих редуцируется правая дуга и сохраняется лишь левая.
Пятая пара дуг у всех позвоночных редуцируется и только у хвостатых амфибий от нее сохраняется незначительный проток. Шестая дуга теряет связь со спинной аортой, от нее берут начало легочные артерии. Сосуд, связывающий во время эмбрионального развития легочную артерию со спинной аортой, называется боталловым протоком. Во взрослом состоянии он сохраняется у хвостатых амфибий и некоторых рептилий. Как порок развития, этот проток может сохраняться и у других более высокоорганизованных животных и человека.
В тесной связи с кровеносной системой находится лимфатическая система: Лимфа выполняет важную роль в обмене веществ, поскольку является посредником между кровью и тканевой жидкостью. Кроме того, она богата лейкоцитами, играющими важную роль в иммунитете.
^ РАЗВИТИЕ СЕРДЦА
В эмбриогенезе человека наблюдается ряд филогенетических преобразований сердца, что имеет важное значение для понимания механизмов развития врожденных пороков сердца.
У низших позвоночных (рыбы, амфибии) сердце закладывается под глоткой в виде полой трубки. У высших позвоночных и человека сердце закладывается в виде двух далеко отстоящих друг от друга трубок. Позже они сближаются, перемещаясь под кишку, а затем смыкаются, образуя единую трубку, расположенную посередине.
У всех позвоночных передняя и задняя части трубки дают начало крупным сосудам. Средняя же часть начинает быстро и неравномерно расти, образуя S-образную форму. После этого задняя часть трубки перемещается на спинную сторону и вперед, образуя предсердие. Передняя часть трубки при этом не смещается, стенки ее утолщаются и она преобразуется в желудочек.
У рыб одно предсердие, а у амфибий оно разделяется растущей перегородкой надвое. Желудочек у рыб и амфибий один, но в желудочке у амфибий имеются мышечные выросты (трабекулы)» образующие мелкие пристеночные камеры. У рептилий в желудочке образуется неполная перегородка, растущая снизу вверх.
У птиц и млекопитающих желудочек разделен на две половины - правую и левую.
В ходе эмбриогенеза у млекопитающих и человека вначале имеется одно предсердие и один желудочек, отделяющиеся друг от друга перехватом с каналом, сообщающим предсердие с желудочком. Затем в предсердии спереди назад начинает расти перегородка, разделяющая предсердие на две части - левое и правое. Одновременно с дорсальной и вентральной сторон начинают расти выросты, которые соединяясь, два отверстия: правое и левое. Позднее в этих отверстиях формируются клапаны. Межжелудочковая перегородка образуется из разных источников.
Н арушение эмбриогенеза сердца может выражаться в отсутствии или неполном заращении межпредсердной или межжелудочковой перегородки. Из аномалий развития сосудов наиболее часто встречается незаращение боталлова протока (от 6 до 22% от всех врожденных пороков сердечно-сосудистой системы), реже - незаращение сонного протока. Кроме того, может вместо одной дуги аорты развиться две - левая и правая, которые образуют аортальное кольцо вокруг трахеи и пищевода с возрастом это кольцо может сужаться и нарушаться глотание. Иногда встречается транспозиция аорты, когда она начинается не от левого желудочка, а от правого, а легочная артерия - от левого.
арушение эмбриогенеза сердца может выражаться в отсутствии или неполном заращении межпредсердной или межжелудочковой перегородки. Из аномалий развития сосудов наиболее часто встречается незаращение боталлова протока (от 6 до 22% от всех врожденных пороков сердечно-сосудистой системы), реже - незаращение сонного протока. Кроме того, может вместо одной дуги аорты развиться две - левая и правая, которые образуют аортальное кольцо вокруг трахеи и пищевода с возрастом это кольцо может сужаться и нарушаться глотание. Иногда встречается транспозиция аорты, когда она начинается не от левого желудочка, а от правого, а легочная артерия - от левого.
^ ЭВОЛЮЦИЯ ЭНДОКРИННОЙ СИСТЕМЫ
Координация работы органов и систем органов у животных обеспечивается наличием двух тесно связанных между собой типов регуляции - нервной и гуморальной. Гуморальная - является более древней и осуществляется через жидкие среды организма с помощью биологически активных веществ, выделяемых клетками и тканями организма в процессе метаболизма
По мере эволюции у животных сформировался специальный аппарат гуморального управления - эндокринная система, или система желез внутренней секреции. С момента появления последней нервная и гуморальная регуляции функционируют в тесной взаимосвязи, образуя единую нейроэндокринную систему.
Гормональная регуляция, в отличие от нервной, направлена, прежде всего, на медленно протекающие реакции в организме, поэтому ей принадлежит главенствующая роль в регуляции формообразовательных процессов: роста, обмена веществ, размножения и дифференцировки.
У беспозвоночных впервые эндокринные железы появляются у кольчатых червей . Наиболее хорошо изучены эндокринные железы у ракообразных и насекомых. Как правило, железы внутренней секреции у этих животных располагаются на переднем конце тела. У ракообразных имеются У-органы, которые вызывают линьку. Эти железы находятся под контролем Х-органов, функционально тесно связанных с головными нервными узлами. Кроме этих желез, у ракообразных в глазных стебельках имеются синусовые железы, регулирующие процессы метаморфоза.
У насекомых на переднем конце тела находятся эндокринные железы, управляющие метаморфозом и стимулирующие энергетический обмен. Эти железы контролируются головной железой внутренней секреции, а последняя - головным нервным узлом. Таким образом, эндокринная система ракообразных напоминает по своей иерархии гипоталамо-гипофизарную систему позвоночных, где гипофиз регулирует работу всех желез внутренней секреции и сам находится под регулирующим влиянием промежуточного мозга.
Э ндокринные железы позвоночных
играют более важную роль в регуляции систем органов, чем у беспозвоночных. У них, кроме шести обособленных желез внутренней секреции (гипофиз, надпочечники, щитовидная железа, паращитовидные железы, тимус, эпифиз), гормоны вырабатываются в раде органов, несущих и другие функции: половые железы, поджелудочная железа, некоторые клетки желудочно-кишечного тракта и др.
ндокринные железы позвоночных
играют более важную роль в регуляции систем органов, чем у беспозвоночных. У них, кроме шести обособленных желез внутренней секреции (гипофиз, надпочечники, щитовидная железа, паращитовидные железы, тимус, эпифиз), гормоны вырабатываются в раде органов, несущих и другие функции: половые железы, поджелудочная железа, некоторые клетки желудочно-кишечного тракта и др.
Эндокринные железы у позвоночных в филогенезе развиваются из разных источников и имеют разное месторасположение. Так щитовидная железа закладывается из эпителия вентральной стороны глотки. У рыб она закладывается между первой и второй жаберными щелями, а у других позвоночных - между вторым и третьим жаберными карманами. Причем, вначале эта железа закладывается как железа внешней секреции. В ходе филогенеза в ряду позвоночных щитовидная железа изменяет свое месторасположение, и, начиная с амфибии, в ней появляются доли и перешеек, что не характерно для рыб, где она имеет вид единого тяжа.
^ Вилочковая железа (тимус ) у рыб развивается за счет эпителиальных выступов, формирующихся на стенках всех жаберных карманов. Эти выступы позже отшнуровываются и образуют две узкие полоски, состоящие из лимфоидной ткани, с просветом внутри.
У амфибий и рептилий количество зачатков, из которых развивается тимус, значительно уменьшается - они берут начало из второй и третьей пары жаберных карманов. У млекопитающих - из трех пар жаберных карманов, но преимущественно из второй пары.
Гипофиз у наземных позвоночных состоит из трех долей: передней, средней (промежуточной) и задней; а у рыб - только из передней и средней.
Гипофиз соединен с нижней поверхностью промежуточного мозга и развивается из разных источников, передняя и средняя доли - из эпителия крыши ротовой полости, а задняя доля - из дистального отдела воронки промежуточного мозга (невральное происхождение). Функция гипофиза у рыб состоит только в выработке гонадотропных гормонов (стимулирующих выработку половыми железами половых гормонов). У амфибий появляется задняя доля, что объясняется их переходом к наземному образу жизни и необходимостью регулировать водный обмен. В заднюю долю заходят аксоны нейросекреторных нейронов гипоталамуса и происходит накопление выделяемого ими антидиуретического гормона с последующим поступлением его в кровь.
Средняя доля, начиная от амфибий, утрачивает способность вырабатывать, гонадотропный гормон, и теперь вырабатывает гормон, стимулирующий синтез меланина. У наземных позвоночных передняя доля, кроме гонадотропина, выделяет и другие тропные гормоны, а также гормон роста.
Надпочечники у хордовых развиваются из двух источников. Их корковое вещество образуется за счет эпителия брюшины, а мозговое имеет невральное происхождение. Причем, у рыб корковое вещество располагается вдоль спинной поверхности первичных почек метамерно и отдельно друг от друга, а мозговое вещество - недалеко от половых валиков по обе стороны от брыжейки
У амфибий между надпочечными телами возникает пространственная связь, а у амниот все закладки надпочечников сливаются, образуя парный орган, состоящий из наружного коркового и внутреннего мозгового, вещества. Надпочечники располагаются над верхним полюсом почек.
^ ЭВОЛЮЦИЯ ИММУННОЙ СИСТЕМЫ
Иммунная система осуществляет защиту организма от проникновения генетически чужеродных тел: микроорганизмов, чужих клеток, инородных тел и др. Ее действие основано на способности отличать собственные структуры организма от генетически чужеродных, элиминируя последние.
В эволюции формировалось три главных формы иммунного ответа: 1) фагоцитоз, или неспецифическое уничтожение генетически чужеродного материала; 2) клеточный иммунитет, основанный на специфическом распознавании его и уничтожении Т-лимфоцитами; 3) гуморальный иммунитет, осуществляемый путем трансформации В-лимфоцитов в плазматические клетки и синтез ими антител (иммуноглобулинов).
В эволюции, выделяют три этапа в формировании иммунного ответа:
- квазиммунное (лат. "квази" - наподобие) распознавание организмом своих и чужеродных клеток. Этот тип реакции наблюдается, начиная от кишечнополостных и кончая млекопитающими. При этом ответе не формируется иммунной памяти, то есть еще не происходит усиления иммунной реакции на повторное проникновение чужеродного материала;
Примитивный клеточный иммунитет обнаружен у кольчатых червей и иглокожих. Он обеспечивается целомоцитами – клетками вторичной полости тела, способными уничтожать чужеродный материал. На этом этапе появляется иммунологическая память;
система интегрированного клеточного и гуморального иммунитета. Для нее характерны специфические гуморальные и клеточные реакции на чужеродные тела, наличие лимфоидных органов иммунитета, образование антител. Такого типа иммунная система не характерна для беспозвоночных.
Тимус, селезенка, отдельные скопления лимфоидной ткани обнаруживаются в полном объеме начиная с амфибий. У низших позвоночных (рыбы, амфибии) вилочковая железа активно выделяет антитела, что не характерно для птиц и млекопитающих.
Особенность иммунной системы иммунного ответа птиц состоит в наличии особого лимфоидного органа - фабрициевой сумки. В этом органе В-лимфоциты после антигенной стимуляции способны трансформироваться в плазматические клетки, вырабатывающие антитела.
У млекопитающих органы иммунной системы разделяют на 2 типа: центральные и периферические. В центральных органах иммуногенеза созревание лимфоцитов происходит без влияния антигенов. В периферических органах иммуногенеза происходит антигензависимое Т и В - размножение и дифференцирование лимфоцитов.
На ранних стадиях эмбриогенеза из желточного мешка в тимус и красный костный мозг мигрируют стволовые лимфатические клетки. После рождения источником стволовых клеток становится красный костный мозг. Периферическими лимфоидными органами являются: лимфоузлы, селезенка, миндалины, лимфоидные фолликулы кишечника. К моменту рождения они еще практически не сформированы и размножение и дифференцировка в них лимфоцитов начинается только после антигенной стимуляции мигрировавших из центральных органов иммуногенеза Т-и В-лимфоцитов.
^ ЭВОЛЮЦИЯ ДЫХАТЕЛЬНОЙ СИСТЕМЫ .
Почти все живые организмы являются аэробами, то есть дышащими воздухом. Совокупность процессов, обеспечивающих поступление и потребление О 2 и выделение СО 2 , называется дыханием.
Функция дыхания у животных разной степени организованности обеспечивается по-разному. Наиболее простой формой дыхания является диффузия газов через стенки живой клетки (у одноклеточных) или через покровы тела (кишечнополостные; плоские, круглые и кольчатые черви). Диффузное дыхание встречается также у мелких членистоногих, имеющих тонкий хитиновый покров и относительно большую поверхность тела.
С усложнением организации животных формируется специальная система органов дыхания; Так уже у некоторых водных кольчецов появляются примитивные органы дыхания - наружные жабры (эпителиальные выросты с капиллярами), При этом в дыхании участвует и кожа. У членистоногих органы дыхания имеют более сложное строение и представлены у водных форм жабрами, а у наземных и вторичноводных легкими и трахеями (у наиболее древних членистоногих, таких как скорпионы, - легкие, у пауков - и легкие и трахеи, а у насекомых, - высших членистоногих, - только трахеи).
Функцию органов дыхания у низших хордовых (ланцетники) берут на себя жаберные щели, по перегородкам которых проходят жаберные артерии (100 пар). Поскольку деление артерий на капилляры в жаберных перегородках отсутствуют, общая поверхность поступления О 2 невелика и окислительные процессы идут на низком уровне. Соответственно этому ланцетник ведет малоподвижный образ жизни.
В связи с переходом позвоночных к активному образу жизни в органах дыхания возникают прогрессивные изменения. Так, у рыб в жаберных лепестках, в отличие от ланцетника, появляется обильная сеть кровеносных капилляров, их дыхательная поверхность резко увеличивается, поэтому число жаберных щелей у рыб сокращается до четырех.
Земноводные - первые животные, вышедшие на сушу, у которых развились органы атмосферного дыхания - легкие (из выпячивания кишечной трубки). В связи с примитивностью строения (легкие представляют собой мешки с тонкими ячеистыми стенками), количество кислорода, поступающего через легкие, удовлетворяет потребность в нем организма только на 30-40%, поэтому в дыхании принимает участие и кожа, содержащая многочисленные кровеносные капилляры (кожно-легочное дыхание).
Воздухоносные пути у амфибий слабо дифференцированы. Они соединены с ротоглоткой небольшой гортано-трахейной камерой.
^
У рептилий
в связи с окончательным выходом на сушу происходит дальнейшее усложнение дыхательной системы: Кожное дыхание исчезает, а дыхательная поверхность легочных мешков увеличивается, благодаря появлению большого количества разветвленных перегородок, в которых проходят кровеносные капилляры. Усложняются и воздухоносные пути: в трахее формируются хрящевые кольца, разделяясь, она дает два бронха. Начинается формирование внутрилегочных бронхов.
рептилий
в связи с окончательным выходом на сушу происходит дальнейшее усложнение дыхательной системы: Кожное дыхание исчезает, а дыхательная поверхность легочных мешков увеличивается, благодаря появлению большого количества разветвленных перегородок, в которых проходят кровеносные капилляры. Усложняются и воздухоносные пути: в трахее формируются хрящевые кольца, разделяясь, она дает два бронха. Начинается формирование внутрилегочных бронхов.
У птиц
в строении органов дыхания появляется ряд особенностей. Легкие у них имеют многочисленные перегородки с сетью кровеносных капилляров. От трахеи идет бронхиальное дерево, заканчивающееся бронхиолами. Часть главных и вторичных бронхов выходит за пределы легких и образует шейные, грудные и брюшные пары воздушных мешков, а также проникает в кости, делая их пневматичными. Во время полета кровь насыщается кислородом и на акте вдоха и на акте выдоха (двойное дыхание).
птиц
в строении органов дыхания появляется ряд особенностей. Легкие у них имеют многочисленные перегородки с сетью кровеносных капилляров. От трахеи идет бронхиальное дерево, заканчивающееся бронхиолами. Часть главных и вторичных бронхов выходит за пределы легких и образует шейные, грудные и брюшные пары воздушных мешков, а также проникает в кости, делая их пневматичными. Во время полета кровь насыщается кислородом и на акте вдоха и на акте выдоха (двойное дыхание).
Млекопитающие имеют легкие альвеолярного строения, благодаря чему их поверхность в 50-100 раз больше поверхности тела. Бронхи древовидно разветвлены и заканчиваются тонкостенными бронхиолами с гроздьями альвеол, густо оплетенных кровеносными капиллярами. Хорошо развиты гортань и трахея.
Таким образом, основное направление эволюции дыхательной системы заключается в увеличении дыхательной поверхности, усложнении строения воздухоносных, путей и их обособлении от респираторных.
^ ЭВОЛЮЦИЯ ВЫДЕЛИТЕЛЬНОЙ СИСТЕМЫ
У одноклеточных животных и кишечнополостных процессы выделения токсических продуктов обмена осуществляются путем их диффузии из клеток во внеклеточную среду. Однако уже у плоских червей появляется система канальцев, выполняющих выделительную и осморегулирующую функции. Эти канальцы называется протонефридиями . Они начинаются крупной звездчатой клеткой, в цитоплазме которой проходит каналец с пучком ресничек, создающих ток жидкости. Эти клетки осуществляют активный транспорт и осмос воды и растворенных вредных веществ в просвет цитоплазматического канальца.
Выделительная система у круглых червей также имеет в своей основе протонефридиальный характер.
У кольчатых червей органами выделения и осморегуляции являются метанефридии . Это канальцы, один конец которых расширен в виде воронки, окружен ресничками и обращен в полость тела, а другой конец открывается на поверхности тела выделительной порой. Выделяемая канальцами жидкость называется мочой. Она образуется путем фильтрации - избирательной реабсорбции и активной секреции из жидкости, содержащейся в полости тела. Метанефридиальный тип выделительной системы характерен и для почек моллюсков.
У членистоногих органами выделения служат либо видоизмененные метанефридии, либо мальпигиевы сосуды, либо специализированные железы
Мальпигиевы сосуды - это пучок трубок, один конец которых слепо заканчивается в полости тела и производит абсорбцию продуктов выделения, а другой открывается в кишечную трубку.
Эволюция выделительной системы хордовых выражается в переходе от нефридий низших хордовых к специальным органам - почкам
У ланцетника выделительная система сходна с таковой у кольчатых червей. Она представлена 100 парами нефридий , один конец которых обращен во вторичную полость тела и всасывает продукты выделения, а другой выводит эти продукты в околожаберную полость.
Органы выделения позвоночных - это парные почки . У низших позвоночных (рыб, амфибий) в эмбриогенезе закладывается два вида почек: предпочка (или головная почка) и туловищная (или первичная). Предпочка напоминает по своему строению метанефридии. Она состоит из извитых канальцев, обращенных воронкой в полость тела, а другим концом впадающих в общий канал предпочки. Недалеко от каждой воронки находится сосудистый клубочек, осуществляющий фильтрацию продуктов обмена в полость тела. Этот вид почки функционирует только в личиночном периоде, а затем начинает функционировать первичная почка. В ней по ходу почечных канальцев имеются выпячивания, в которых располагаются сосудистые клубочки и происходит фильтрация мочи. Воронки утрачивают свое функциональное значение и зарастают.
У высших позвоночных в эмбриональном периоде закладывается последовательно три почки: предпочка, первичная (туловищная) и вторичная (тазовая) почка . Предпочка не функционирует. Первичная почка функционирует только в эмбриогенезе. Ее проток расщепляется на два: вольфов и мюллеров каналы. В дальнейшем вольфовы каналы преобразуются в мочеточники, а у самцов в мочеточники и семяпроводы. Мюллеровы каналы сохраняются только у самок, преобразуясь в яйцеводы. Т.е. в эмбриогенезе мочевая и половая системы связаны.
К концу эмбрионального периода начинает функционировать тазовая (вторичная) почка. Это компактные парные образования, расположенные по бокам поясничного отдела позвоночника. Морфо-функциональной единицей в них являются нефроны, которые состоят из капсулы с сосудистым клубочком системы извитых канальцев первого и второго порядка и петли Генле. Канальцы нефрона переходят в собирательные трубки, которые открываются в почечные лоханки.
^ ЭВОЛЮЦИЯ ИММУННОЙ СИСТЕМЫ
Иммунная система осуществляет защиту организма от проникновения в организм генетически чужеродных тел: микроорганизмов, вирусов, чужих клеток, инородных тел. Ее действие основано на способности отличать собственные структуры от генетически чужеродных, элиминируя их.
В эволюции сформировалось три главных формы иммунного ответа:
фагоцитоз - или неспецифическое уничтожение генетически чужеродного
клеточный иммунитет, основанный на специфическом распознавании и уничтожении такого материала Т-лимфоцитами;
гуморальный иммунитет, осуществляемый путем образования потомками В-лимфоцитов, так называемыми, плазматическими клетками иммуноглобулинов и связывания ими чужеродных антигенов.
I этап - квазииммунное (лат. квази - наподобие, как бы) распознавание организмом своих и чужеродных клеток. Этот тип реакции наблюдается от кишечнополостных до млекопитающих. Эта реакция не связана с выработкой иммунных тел, и при этом не формируется иммунной памяти, то есть еще не происходит усиления иммунной реакции на повторное проникновение чужеродного материала.
II этап - примитивный клеточный иммунитет обнаружен у кольчатых червей и иглокожих. Он обеспечивается целомоцитами - клетками вторичной полости тела, способными уничтожать чужеродный материал. На этом этапе появляется иммунологическая память.
III этап - система интегрированного клеточного и гуморальго иммунитета. Для нее характерны специфические гуморальные и клеточные реакции на чужеродные тела. Характерно наличие лимфоидных органов иммунитета, образование антител. Такого типа иммунная система не характерна для беспозвоночных.
Круглоротые способны формировать антитела, но вопрос о наличии у них вилочковой железы, как центрального органа иммуногенеза, является пока открытым. Впервые тимус обнаруживается у рыб.
Эволюционные предшественники лимфоидных органов млекопитающих - тимус, селезенка, скопления лимфоидной ткани обнаруживаются в полном объеме у амфибий . У низших позвоночных (рыбы, амфибии) вилочковая железа активно выделяет антитела, что не характерно для птиц и млекопитающих.
Особенность системы иммунного ответа птиц состоит в наличии особого лимфоидного органа - фабрициевой сумки . В этом органе образуются В-лимфоциты, которые после антигенной стимуляции способны трансформироваться в плазматические клетки и вырабатывать антитела.
У млекопитающих органы иммунной системы разделяют на два типа: центральные и периферические. В центральных органах созревание лимфоцитов происходит без существенного влияния антигенов. Развитие периферических органов, наоборот, непосредственно зависит от антигенного воздействия - лишь при контакте с антигеном в них начинаются процессы размножения и дифференциации лимфоцитов.
Центральным органом иммуногенеза у млекопитающих является тимус, где образуются Т-лимфоциты, а также красный костный мозг, где образуются В-лимфоциты.
На ранних стадиях эмбриогенеза из желточного мешка в тимус и красный костный мозг мигрируют стволовые лимфотические клетки. После рождения источником стволовых клеток становится красный костный мозг.
Периферическими лимфоидными органами являются: лимфоузлы, селезенка, миндалины, лимфоидные фолликулы кишечника. К моменту рождения они еще не сформированы практически и образование в них лимфоцитов начинается только после антигенной стимуляции, после того, как они заселяются Т-и В-лимфоцитами из центральных органов иммуногенеза.
^ ФИЛОГЕНЕЗ ВИСЦЕРИАЛЬНОГО ЧЕРЕПА У ПОЗВОНОЧНЫХ ЖИВОТНЫХ .
Череп позвоночных состоит из двух основных отделов - осевого и висцерального.
Осевой - черепная коробка (мозговой череп - neurocranium) - продолжение осевого скелета, служит для защиты головного мозга и органов чувств.
Висцеральный - лицевой (splanchnocranium), образует опору для передней части пищеварительного тракта.
 ба отдела черепа развиваются независимо друг от друга, разными путями. Висцеральная часть черепа у зародышей позвоночных животных состоит из метамерно расположенных хрящевых дуг, которые охватывают переднюю часть пищеварительного тракта, и отделены друг от друга висцеральными щелями. Дуги обозначают порядковыми номерами в соответствии с расположением по отношению к черепу.
ба отдела черепа развиваются независимо друг от друга, разными путями. Висцеральная часть черепа у зародышей позвоночных животных состоит из метамерно расположенных хрящевых дуг, которые охватывают переднюю часть пищеварительного тракта, и отделены друг от друга висцеральными щелями. Дуги обозначают порядковыми номерами в соответствии с расположением по отношению к черепу.Первая дуга у большинства современных позвоночных приобретает функцию челюстного аппарата - ее называют челюстной, а вторую - тоже по функции - подъязычной или гиоидной. Остальные, начиная с третьей и до седьмой, называют жаберными, т.к. они служат опорой для жаберного аппарата. На ранних этапах развития висцеральный и осевой череп не связаны друг с другом, позднее эта связь возникает.
Общие для всех зародышей позвоночных закладки семи висцеральных дуг в процессе эмбрионального развития претерпевают у представителей разных классов соответственно и различные специфические изменения.
^ I. Низшие рыбы (хрящевые) - Chondrichthyes
1-я, она же челюстная дуга, состоит из двух крупных хрящей, вытянутых в переднезаднем направлении: верхнего - небноквадратного - первичная верхняя челюсть, нижнего - меккелева - первичная нижняя челюсть; они сзади сращены между собой и выполняют функцию первичной челюсти.
2-я, она же подъязычная, или гиоидная дуга состоит из следующих компонентов:
из двух, расположенных, вверху гиомандибулярных хрящей, которые сверху соединены с черепной коробкой, снизу- с гиоидом, а впереди - с челюстной дугой - первичной верхней челюстью;
из двух гиоидов, расположенных ниже гиомандибулярных хрящей, которые с ними и соединены; кроме того, гиоиды соединены с первичной нижней челюстью;
из непарной копулы (небольшой хрящ, соединяющий оба гиоида между собой).
Остальные висцеральные дуги с третьей по седьмую образуют опору для дыхательного аппарата.
^ II.Высшие рыбы - (костные) Osteichthyes .
Основное отличие касается только челюстной дуги:
1) верхний элемент челюстной дуги (верхняя челюсть) состоит вместо одного крупного небноквадратного хряща из пяти элементов - небного хряща, квадратной кости и трех крыловидных хрящей;
впереди от первичной верхней челюсти образуются две крупные накладные кости, снабженные крупными зубами, - эти кости и становится - вторичными верхними челюстями;
дистальный конец первичной нижней челюсти тоже покрывает большая зубная кость, которая выдается далеко вперед и образует вторичную нижнюю челюсть. Подъязычная дуга сохраняет прежнюю функцию, т.е. череп остается гипостильным.
Главное отличие в новом способе соединения челюстной дуги с черепом: небный хрящ первичной верхней челюсти на всем протяжении срастается с осевым черепом, т.е. с черепной коробкой. Такой тип соединения называется аутостильный.
Нижнечелюстной отдел соединяется с верхнечелюстным и тоже получает связь с черепом без подъязычной дуги.
Таким образом, гиомандибулярный хрящ освобождается от функции подвеска, существенно редуцируется и приобретает новую функцию - входит в состав воздухоносной полости среднего уха в виде слуховой косточки - столбика.
Часть подъязычной дуги (гиоидный хрящ), жаберные дуги образуют частичную опору для языка и подъязычный аппарат, частично гортанные хрящи, частично редуцируются.
^ IV.Пресмыкающиеся - Reptilia .
Череп аутостильный, но при этом небный хрящ первичной челюсти редуцируется и в причленении верхней челюсти к черепу участвует только квадратная кость, с ней соединяется нижняя челюсть и таким образом присоединяется к черепу. Остальная часть висцерального скелета образует подъязычный аппарат, который состоит из тела подъязычной кости и трех пар отростков.
^ V.Млекопитающие - Mammalia .
Появляется совершенно новый способ соединения с черепом нижней челюсти, которая присоединяется к нему непосредственно, образуя сустав с чешуйчатой костью черепа, что позволяет не только захватывать пищу, но и совершать сложные жевательные движения. В образовании сустава участвует только вторичная нижняя челюсть. Следовательно, квадратная кость первичной верхней челюсти теряет свою функцию подвеска и превращается в слуховую косточку - наковальню.
Первичная нижняя челюсть в процессе эмбрионального развития полностью выходит из состава нижней челюсти и преобразуется в следующую слуховую косточку - молоточек.
Верхний отдел подъязычной дуги, гомолог гиомандибулярного хряща преобразуется в стремечко.
Все три слуховые косточки составляют единую функциональную цепь.
1-я – жаберная дуга (1-я висцеральная) и копула дают, начало телу подъязычной кости и ее задними рожкам.
2-я и 3-я жаберные дуги (4-я и 5-я висцеральные) дают начало щитовидному хрящу, который впервые появляется у млекопитающих.
4-я и 5-я жаберные дуги (1-я и 7-я висцеральные) дают материал для остальных гортанных хрящей, и возможно, для трахеальных.
^ ЭВОЛЮЦИЯ ЗУБНОЙ СИСТЕМЫ
И РОТОВЫХ ЖЕЛЕЗ ПОЗВОНОЧНЫХ
Рыбы-Piscеs
Зубная система - гомодонтная (зубы одинаковые). Зубы имеют коническую форму, обращены назад, служат для удержания пищи, располагаются по краю черепа, у некоторых на всей поверхности ротовой полости.
Слюнных желез в ротовой полости нет, т.к. они пищу заглатывают с водой. Язык примитивный, в виде двойной складки слизистой оболочки. Крыша ротовой полости образована основанием мозгового черепа - первичное твердое небо. Ротовое отверстие окружено кожными складками - губами, которые неподвижны. Общая ротоглоточная полость.
Плакоидная чешуя хрящевых рыб представляет собой пластинку с положенным на ней шипом. Пластинка лежит в кориуме вершина шипа через эпидермис выдается наружу. Вся чешуя состоит из дентина, образованного клетками кориума, вершина шипа покрыта эмалью, образуемой клетками базального слоя эпидермиса.
Более крупные и усложненные плакоидные чешуи располагаются в челюстях, образуя зубы. В сущности, зубы всех позвоночных - это измененные плакоидные чешуи предков.
Земноводные - Amphibia.
Зубная система гомодонтная. Зубы у ряда земноводных располагаются не только на альвеолярной дуге им, как и рыбам, свойствен полифиодонтизм.
Появляются слюнные железы, секрет которых не содержит ферментов. В языке располагаются мышцы, определяющие его собственную подвижность. Крыша ротовой полости - тоже первичное твердое небо. Губы неподвижны Общая ротоглоточная полость.
Пресмыкающиеся - Reptilia.
Зубная система у современных рептилий гомодонтная , ядовитые пресмыкающиеся имеют специальные зубы, по которым яд стекает в ранку от укуса. Зубы располагаются в один ряд. У некоторых вымерших форм обнаруживается начальная дифференцировка. Всем рептилиям свойствен полифиодонтизм.
Слюнные железы развиты лучше, среди них выделяют подъязычные, зубные и губные. Секрет желез уже содержит ферменты.
У ядовитых змей задняя пара зубных желез преобразована в ядовитые, секрет содержит токсины (яд).
Язык закладывается из трех зачатков: один - непарный и два - парных, лежащих впереди непарного. Парные зачатки позже срастастаются. У большинства рептилий это срастание неполное и язык раздвоен.
Возникают зачатки вторичного твердого неба в виде горизонтальных костных складок верхней челюсти, которые доходят до середины и делят ротовую полость на верхний отдел - дыхательный (носоглоточный) и нижний - вторичную ротовую полость. Губы неподвижны.
Млекопитающие - Mammalia,
Зубы гетеродонтные , т.е. дифференцированные: различают резцы (incisivi), клыки (canini), малые коренные (praemolares) и коренные (molares). У ластоногих и зубатых китов зубы не дифференцированы. Зубы сидят в альвеолах, на альвеолярных дугах челюстей основание зуба сужается, образуя корень.
Резцы и клыки весьма сходны с коническими зубами предков (рептилий), коренные зубы подверглись наибольшим эволюционным преобразованиям и впервые возникли у зверозубых ящеров.
В связи с дифференцировкой зубов увеличивается продолжительность функционирования. В онтогенезе существует две смены зубов (дифиодонтизм
): резцы, клыки и большие коренные имеют две генерации (молочные и постоянные); малые коренные - только одну.
связи с дифференцировкой зубов увеличивается продолжительность функционирования. В онтогенезе существует две смены зубов (дифиодонтизм
): резцы, клыки и большие коренные имеют две генерации (молочные и постоянные); малые коренные - только одну.
Общее количество зубов у разных отрядов – разное: например, у слонов - 6, волка - 42, кошек - 30, зайцев - 28, у большинства приматов и человека-32.
Слюнные железы млекопитающих многочисленны: это и мелкие - язычные, щечные, небные, зубные - гомологичные железам рептилий, и крупные - подъязычные, подчелюстные, околоушные. Из них первые две появились в результате дифференцировки подъязычной железы рептилий, а околоушные - новое приобретение млекопитающих. В ротовой полости - у высших млекопитающих появляются крупные скопления лимфатической ткани - миндалины.
Язык также, как и у пресмыкающихся, развивается из трех зачатков. Вторичное твердое небо становится сплошным, полностью отделяется полость рта от полости носа, чем достигается независимость функций ротовой полости и дыхания. Кзади твердое небо продолжается в мягкое - двойная складка слизистой, отделяющая ротовую полость глотки. Поперечные валики твердого неба способствуют перетиранию пищи. У человека они постепенно исчезают после рождения.
Губы мясистые у сумчатых и плацентарных, подвижны, что связано со вскармливанием детенышей молоком. Губы, щеки и челюсти ограничивают пространство, называемое преддверием рта.
^ У человека зубная формула 2123
2123(половина верхней и нижней челюсти).
Зубы, по сравнению с другими приматами, уменьшились в размерах, особенно клыки, они не выступают из зубного ряда и не перекрываются. Исчезли диастемы (промежутки между зубами) в верхних и нижних челюстях, зубы стали в плотный ряд, зубная дуга получила округлую (параболическую) форму.
М оляры имеют четырехбугорчатую форму. Последняя пара моляров, "зубы мудрости", прорезываются поздно - до 25 лет. Они явно рудиментарны, уменьшены в размерах и часто слабо дифференцированы.
оляры имеют четырехбугорчатую форму. Последняя пара моляров, "зубы мудрости", прорезываются поздно - до 25 лет. Они явно рудиментарны, уменьшены в размерах и часто слабо дифференцированы.
Во время жевания нижняя челюсть может совершать вращательные движения по отношению к верхней, благодаря неперекрыванию редуцированных клыков и комплементарно расположенным буграм жевательных зубов обеих челюстей.
^ АТАВИСТИЧЕСКИЕ АНОМАЛИИ РОТОВОЙ ПОЛОСТИ ЧЕЛОВЕКА:
А) редкая аномалия - гомодонтная зубная система, все зубы конической формы;
Б) трехбугорчатые коренные зубы;
В) прорезывание сверхкомплектных зубов, т.е. у человек возможно образование большего, чем 32, количества зубных зачатков;
Г) отсутствие "зубов мудрости";
Д) весьма редкий порок развития языка - раздвоенность его конца, как результат несращения парных зачатков в эмбриогенезе;
Е) нарушение срастания (оно должно произойти к концу восьмой недели эмбриогенеза) костных горизонтальных складок которые образуют твердое небо, приводит к незаращению твердого неба и формированию порока, известного под названием "волчья пасть";
Ж) расщелина верхней губы ("заячья губа") возникает за счет неполного срастания формирующих верхнюю губу кожно-мезодермальный выростов, два из которых (латеральные) растут от верхней челюсти, а один (центральный) - от лобно-носового отростка.
^ СИНТЕТИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИ
Начавшееся в 20-х годах объединение дарвинизма с экологией и генетикой подготовило почву для создания синтетической теории эволюции, на сегодняшний день единственно целостной, достаточно полно разработанной теорией биологической эволюции, воплотившей классический дарвинизм и популяционную генетику.
Первым ученым, который ввел генетический подход в изучение эволюционных процессов был Сергей Сергеевич Четвериков. В 1926 г - он опубликовал научную статью "О некоторых моментах эволюционного процесса с точки зрения современной генетики", в которой ему удалось показать на примере природных популяций дрозофил, что: 1) в природных популяциях постоянно происходят мутации; 2) рецессивные мутации, "как губкой впитываются" видом и в гетерозиготном состоянии могут сохраняться неограниченно долго; 3) по мере старения вида в нем накапливается все больше мутаций и признаки вида при этом расшатываются; 4) изоляция и наследственная изменчивость являются основными факторами внутривидовой дифференцировки; 5) панмиксия ведет к полиморфизму вида, а отбор - к мономорфизму. В этой работе С.С Четвериков подчеркивает, что накопления отбором малых случайных мутаций приводит к закономерному, адаптивно направленному течению эволюции. Работы С.С. Четверикова были продолжены такими отечественными генетиками, как Н.В. Тимофеев-Ресовский, Д.Д. Ромашов. Н.П. Дубинин, Н.И Вавилов и др. Эти работы подготовили почву для создания основ синтетической теории эволюции.
В 30-х годах работами английских ученых Р. Фишера. Дж. Холдейма. С. Райта было положено начало синтеза теории эволюции и генетики на Западе.
Одной из первых работ, изложивший суть синтетической теории эволюции" была монография Ф.Г. Добжанского "Генетика и происхождение видов" (1937 г.). Основное внимание в этой работе было обращено на изучение механизмов формирования генетической структуры популяций в зависимости от воздействия таких факторов и причин эволюции, как наследственная изменчивость, естественный отбор, колебания численности особей в популяциях (популяционные волны), миграция и, наконец, репродуктивная изоляция новых форм, возникших внутри вида.
Выдающийся вклад в создание синтетической теории эволюции внес отечественный ученый И.И. Шмальгаузен. На основе творческого объединения эволюционной теории, эмбриологии, морфологии, палеонтологии и генетики он глубоко исследовал соотношение онтогенеза и филогенеза, изучил основные направления эволюционного процесса, разработал ряд фундаментальных положений современной теории эволюции. Его основные работы: "Организм как целое в индивидуальном и историческом развитии" (1938); "Пути и закономерности эволюционного процесса" (1939); "Факторы эволюции" (1946).
Важное место среди фундаментальных исследований по теории эволюции занимает вышедшая в 1942 году под редакцией видного английского эволюциониста Джулиана Хаксли монография "Эволюция. Современный синтез" (1942), а также исследования темпов и форм эволюции предпринятые Джорджем Симпсоном,
В основе синтетической теории эволюции лежат 11 основных постулатов, которые в сжатой форме сформулированы отечественным современным генетиком Н.Н. Воронцовым, примерно, таком виде:
1. Материалом для эволюции служат, как правило, очень мелкие, дискретные изменения наследственности – мутации. Мутационная изменчивость как поставщик материала для отбора носит случайный характер. Отсюда название концепции, предложенной ее критиком Л.С. Бергом (1922), "тихогенез", эволюция на основе случайностей.
Основным или даже единственным движущим фактором эволюции является естественный отбор, основанный на отборе (селекции) случайных и мелких мутаций. Отсюда название теории – селектогенез.
Наименьшая эволюционирующая единица - это популяция, а не особь, как допускалось Ч. Дарвином. Отсюда особое внимание к изучению популяции как структурной единицы сообществ: вида, стада, стаи.
Эволюция носит постепенный (градационный) и длительный характер. Видообразование мыслится как поэтапная смена одной временной популяции чередой последующих временных популяций.
Вид состоит из множества соподчиненных, в то же время морфологически, физиологически и генетически отличимых, но репродуктивно не изолированных, единиц - подвидов, популяций (концепция широкого политипического вида).
Эволюция носит дивергентный характер (расхождение признаков), т.е. один таксон (систематическая группировка) может стать предком нескольких дочерних таксонов, но каждый вид имеет один единственный предковый вид, единственную предковую популяцию.
Обмен аллелями (поток генов) возможен лишь внутри вида. Отсюда вид является генетически замкнутой и целостной системой.
Критерии вида не применимы к формам, размножающимся бесполым путем и партеногенетически. Таковыми могут быть огромное множество прокариот, низших эукариот без полового процесса, а также некоторые специализированные формы высших эукариот, вторично утратившие половой процесс (размножаются партеногенетически)
Макроэволюция (т.е. эволюция выше вида) идет путем микроэволюции.
11. Эволюция непредсказуема, т.е., имеет ненаправленный к конечной цели характер.
В конце 50-х начале 60-х годов XX века появляются дополнительные сведения, указывающие на необходимость пересмотра некоторых положений синтетической теории. Назревает необходимость в коррекции ее некоторых положений.
В настоящее время к 1,2 и 3-й тезисы теории остаются в силе:
4-й тезис считается необязательным, т.к. эволюция иногда может идти очень быстро скачками. В 1982 году в г. Дижоне (Франция) состоялся симпозиум, посвященный вопросам темпов и формам видообразования. Было показано, что в случае полиплоидиии хромосомных перестроек, когда практически сразу формируется репродуктивная изоляция, видообразование идет скачкообразно. Тем не менее, в природе без сомнения существует и постепенное видообразование путем отбора мелких мутаций.
5-й постулат оспаривается, поскольку известно немало видов с ограниченным ареалом, в пределах которого не удается их расчленить на самостоятельные подвиды, а реликтовые виды вообще могут состоять из одной популяции, и судьба таких видов, как правило, недолговечна.
7-й тезис в основном остается в силе. Однако, известны случаи просачивания генов через барьеры изолирующих механизмов между особями разных видов. Существует, так называемый, горизонтальный перенос генов, например, трансдукция - перенос бактериальных генов от одного вида бактерий к другим через инфицирование их бактериофагами. Вокруг вопроса о горизонтальном переносе генов идут дискуссии. Число публикаций по этому вопросу растет. Новейшая сводка представлена в монографии Р.Б. Хасина "Непостоянство генома" (1984).
Подлежат также рассмотрению с эволюционных позиций транспозоны, которые, мигрируя внутри генома, приводят к перераспределению последовательности включения тех или иных генов.
8-й тезис требует доработки, так как непонятно, куда включать организмы, размножающиеся неполовым путем, которые по этому критерию не могут, быть отнесены к определенным видам.
9-й тезис в настоящее время пересматривается, поскольку имеются сведения о том, что макроэволюция может идти как через микроэволюцию, так и минуя традиционные микроэволюционные пути.
10-й тезис - возможность дивергентного происхождения таксонов от одной предковой популяции (или вида) ныне никем не отрицается. Но эволюция не всегда носит дивергентный характер. В природе распространена также форма происхождения новых таксонов путем слияния разных, ранее независимых, т.е. репродуктивно изолированных, ветвей. Объединение разных геномов и создание нового сбалансированного генома идет на фоне действия естественного отбора, отбрасывающего нежизнеспособные комбинации геномов. В 30-х годах учеником Н.И. Вавилова В.И. Рыбиным был осуществлен ресинтез (обратный синтез) культурной сливы, происхождение которой не было выяснено. В.А. Рыбин создал ее копию путем гибридизации терна и алычи. Ресинтезом было доказано гибридогенное происхождение некоторых других видов дикорастущих растений. Ботаники считают гибридизацию одним из важных путей эволюции растений.
11-й тезис также пересматривается. Особое внимание эта проблема начала привлекать в начале 20-х годов нашего столетия, когда появились работы Н.И. Вавилова о гомологических рядах наследственной изменчивости. Он обратил внимание на существование некоторой направленности в изменчивости организмов и высказал мысль о возможности прогнозирования ее на основании анализа рядов гомологичной изменчивости у родственных форм организмов.
В 20-х годах появились работы отечественного ученого Л.А. Берга, который высказал мысль о том, что эволюция носит в некоторой степени предопределенный, канализированный характер, что существуют некие запретные пути эволюции, поскольку число оптимальных решений в ходе этого процесса, видимо, бывает ограниченным (теория номогенеза).
Исходя из современных представлений, можно сказать, что в эволюции существует определенная векторизованность путей преобразования признаков, и мы можем в какой-то степени предсказать направленность эволюции.
Итак, современная теория эволюции накопила огромный арсенал новых фактов и идей, однако целостной теории, могущей заменить синтетическую теорию эволюции, пока нет и это дело будущего.
После выхода в свет основного труда Ч. Дарвина "Происхождение видов путем естественного отбора" (1859) современная биология далеко отошла не только от классического дарвинизма второй половины ХIX века, но и от ряда положений синтетической теории эволюции. Вместе с тем несомненно, что магистральный путь развития эволюционной биологии лежит в русле тех направлений, которые были заложены Дарвином.
^ ГЕНЕТИЧЕСКИЙ ПОЛИМОРФИЗМ
Под генетическим полиморфизмом понимается состояние длительного разнообразия генотипов, когда частота даже наиболее редко встречающихся генотипов в популяциях превышают 1%. Генетический полиморфизм поддерживается за счет мутаций и рекомбинаций генетического материала. Как показывают многочисленные исследования, генетический полиморфизм широко распространен. Так, по теоретическим расчетам в потомстве от скрещивания двух особей, различающихся лишь по десяти локусам, каждый из которых представлен 4 возможными аллелями, окажется около 10 млрд. особей с различными генотипами.
Чем больше запас генетического полиморфизма в данной популяции, тем легче ей адаптироваться к новой среде и тем быстрее протекает эволюция. Однако, оценить количество полиморфных аллелей посредством традиционных генетических методов практически невозможно, поскольку сам факт присутствия какого-либо гена в генотипе устанавливается путем скрещивания особей, обладающих различными формами фенотипа, определяемого этим геном. Зная, какую долю в популяции составляют особи с различными фенотипами, можно выяснить, сколько аллелей участвуют в формировании данного признака.
Начиная с 60-х годов XX столетия для определения генетического полиморфизма стал широко применяться метод электрофореза белков (в том числе и ферментов) в геле. С помощью этого метода можно вызвать перемещение белков в электрическом поле в зависимости от их размера, конфигурации и суммарного заряда в разные участки геля, а затем по расположению и числу проявляющихся при этом пятен проводить идентификацию исследуемого вещества. Для оценки степени полиморфизма тех или иных белков в популяциях обычно исследуют около 20 и более локусов, а потом математическим путем определяют количество аллельных генов, соотношение гомо - и гетерозигот. Как показывают исследования, одни гены, как правило, бывают мономорфными, а другие - чрезвычайно полиморфными.
Различают переходный и сбалансированный полиморфизм, что зависит от селективной ценности генов и давления естественного отбора.
^ Переходный полиморфизм возникает в популяции, когда происходит замещение аллеля, бывшего некогда обычным, другими аллелями, придающими своим носителям более высокую приспособленность (множественный аллелизм). При переходном полиморфизме наблюдается направленный сдвиг в процентном соотношении форм генотипов. Переходный полиморфизм - это главный путь эволюции, ее динамика. Примером переходного полиморфизма может быть явление индустриального механизма. Так, в результате загрязнения атмосферы в промышленных городах Англии за последние сто лет у более чем 80 видов бабочек, появились темные формы. Например, если до 1848 г. березовые пяденицы имели бледно-кремовую окраску с черными точками и отдельными темными, пятнами, то в 1848 г. в Манчестере появились первые темнотелые формы, а к 1895 г. уже 98% пядениц стало темнотелыми. Это произошло вследствие закопчения стволов деревьев и избирательного выедания светлотелых пядениц дроздами и малиновками. Позже было установлено, что темная окраска тела у пядениц осуществляется мутантным меланистическим аллелем.
^ Сбалансированный полиморфизм х арактеризуется отсутствием сдвига числовых соотношений различных форм, генотипов в популяциях, находящихся в стабильных условиях среды. При этом процентное соотношение форм либо из поколения в поколение остается одним и тем же, либо колеблется вокруг какой-то постоянной величины. В противоположность переходному, сбалансированный полиморфизм - это статика эволюции. И.И. Шмальгаузен (1940) назвал его равновесным гетероморфизмом.
Примером сбалансированного полиморфизма служит наличие двух полов у моногамных животных, поскольку они обладают равноценными селективными преимуществами. Их соотношение в популяциях составляет 1:1. При полигамии селективное значение у представителей разных полов может отличаться и тогда представители одного пола либо уничтожаются, либо в большей степени, чем особи другого пола, отстраняются от размножения. Другой пример - группы крови человека по АВО-системе. Здесь частота разных генотипов в различных популяциях может варьировать, однако, в каждой конкретной популяции она остается постоянной из поколения в поколение. Это объясняется тем, что ни один генотип не обладает селективным преимуществом перед другими. Так, хотя у мужчин с первой группы крови, как показывает статистка, ожидаемая продолжительность жизни, выше, чем у мужчин с другими группами крови, у них чаше, чем у других, развивается язва двенадцатиперстной кишки, которая в случае прободения может привести к смерти.
Генетическое равновесие в популяциях может нарушаться давлением спонтанных мутаций, возникающих с определенной частотой в каждом поколении. Сохранение или же элиминация этих мутаций зависит от того, благоприятствует ли им естественный отбор или противодействует. Прослеживая судьбу мутаций в той или иной популяции, можно говорить о ее адаптивной ценности. Последняя равна 1, если отбор не исключает ее и не противодействует распространению. В большинстве случаев показатель адаптивной ценности мутантных генов оказывается меньше 1, а если мутанты совершенно не способны размножаться, то он равен нулю. Такого рода мутации отметаются естественным отбором. Однако, один и тот же ген может неоднократно мутировать, что компенсирует его элиминацию, производимую отбором. В таких случаях может быть достигнуто равновесие, когда появление и исчезновение мутировавших генов становится сбалансированным. Примером может служить серповидноклеточная анемия, когда доминантный мутантный ген в гомозиготе приводит к ранней гибели организма, однако, гетерозиготы по этому гену оказываются устойчивыми к заболеванию малярией. В районах, где распространена малярия, имеет место сбалансированный полиморфизм по гену серповидноклеточной анемии, поскольку наряду с элиминацией гомозигот, действует контротбор в пользу гетерозигот. В результате разновекторного отбора в генофонде популяций поддерживаются в каждом поколении генотипы, обеспечивающие приспособленность организмов с учетом местных условий. Помимо гена в серповидноклеточности, в популяциях человека есть ряд других полиморфных генов, которые как предполагают, вызывают явление гетерозиса
Рецессивные мутации (в том числе и вредные), не проявляющиеся фенотипически у гетерозигот, могут накапливаться в популяциях до более высокого уровня, чем вредные доминантные мутации.
Генетический полиморфизм является обязательным условием для непрерывной эволюции. Благодаря ему в изменяющейся среде всегда могут быть генетические варианты предаптированные к этим условиям. В популяции диплоидных раздельнополых организмов может храниться в гетерозиготном состоянии, не проявляясь фенотипически, огромный запас генетической изменчивости. Уровень последней, очевидно, может быть еще более высоким у полиплоидных организмов, у которых за фенотипически проявляющимся нормальным аллелем может скрываться не один, а несколько мутантных аллелей.
^ ГЕНЕТИЧЕСКИЙ ГРУЗ
Генетическая гибкость (или пластичность) популяций достигается за счет мутационного процесса и комбинативной изменчивости. И хотя эволюция зависит от постоянного наличия генетической изменчивости, одно из ее последствий - это появление в популяциях слабо адаптированных особей, в результате чего приспособленность популяций всегда оказывается ниже той, которая характерна для оптимально приспособленных организмов. Это снижение средней приспособленности популяции за счет особей, приспособленность которых ниже оптимальной, называют генетическим грузом . Как писал известный английский генетик Дж. Холдейн, характеризуя генетический груз: "Это та цена, которую вынуждена платить популяция за право эволюционировать". Он был первым, кто привлек внимание исследователей к существованию генетического груза, а сам термин "генетический груз" ввел в 40-х годах XX века Г. Миллер.
Генетический груз в его широком смысле - это всякое снижение (действительное или потенциальное) приспособленности популяции в силу генетической изменчивости. Дать количественную оценку генетического груза, определить его подлинное влияние на популяционную приспособленность - сложная задача. По предложению Ф. Г. Добжанского (1965) носителями генетического груза считаются индивидуумы, приспособленность которых более чем на два стандартных отклонения (-2а) ниже средней приспособленности гетерозигот.
Принято выделять три вида генетического груза: мутационный, субстиционный (переходный) и сбалансированный. Общий генетический груз слагается из этих трех видов груза. Мутационный груз - это та доля общего генетического груза, которая возникает за счет мутаций. Однако, поскольку большинство мутаций носят вредный характер, то естественный отбор направлен против таких аллелей и частота их невелика. Они поддерживаются в популяциях в основном благодаря вновь возникающим мутациям и гетерозиготным носителям.
Генетический груз, возникающий при динамическом изменении частот генов в популяции в процессе замены одного аллеля другим, называется субстиционный (или переходным) грузом . Такое замещение аллелей обычно происходит в ответ на какое-либо изменение в условиях среды, когда ранее неблагоприятные аллели становятся благоприятными, и, наоборот, (примером может быть явление индустриального механизма бабочек в экологически неблагополучных районах). При этом частота одного аллеля уменьшается по мере увеличения частоты другого.
^ Сбалансированный (устойчивый) полиморфизм возникает, когда многие признаки поддерживаются на относительно постоянном уровне за счет уравновешивающего отбора. При этом благодаря сбалансированному (уравновешивающему) отбору, действующему в противоположных направлениях в популяциях сохраняются два или больше аллея ей какого-либо локуса, а соответственно и разные генотип и фенотипы. Примером может служить серповидноклеточность. Здесь отбор направлен против мутантного аллеля, находящегося в гомозиготном состоянии, но в то же время действует в пользу гетерозигот, сохраняя его. Состояние сбалансированного груза может быть достигнуто в следующих ситуациях: 1) отбор благоприятствует данному аллелю на одной стадии онтогенеза и направлен против него на другой; 2) отбор благоприятствует сохранению аллеля у особей одного пола и действует против - у особей другого пола; 3) в пределах одного аллеля разные генотипы дают возможность организмам использовать разные экологические ниши, что снижает конкуренцию и, как следствие, ослабляется элиминация; 4) в субпопуляциях, занимающих разные места обитания, отбор благоприятствует разным аллелям; 5) отбор благоприятствует сохранению аллеля пока он редко встречается и направлен против него, когда он встречается часто.
Предпринималось много попыток оценить реальный генетический груз в популяциях человека, однако, оказалось, что это очень сложная задача. Косвенно о нем можно судить по уровню пренатальной смертности и рождению детей с теми или иными формами аномалий развития, особенно от родителей, состоящих в инбредных браках, а еще более – инцестных.
Литература:
1.Абрикосов Г.Г., Беккер З.Г. и др. Курс зоологии в двух томах. Том I.- Зоология беспозвоночных. Издание 7-е. Изд.: «Высшая школа», М.,1966.-552с.
2.Бакл, Джон. Гормоны животных (пер. с англ. М.С. Морозовой). Изд.:Мир, 1986.-85(1)с.
3.Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. Изд.:Сов. Наука, М., 1944.-489с.
4.Волкова О.В., Пекарский М.И. Эмбриогенез и возрастная гистология внутренних органов человека. Изд.: «Медицина», М,1976. 45с.
5.Гуртовой Н.Н., Матвеев Б.С., Дзержинский Ф.Я. Практическая зоотомия позвоночных. Земноводные и пресмыкающиеся./Под ред. Б.С. Матвеева и Н.Н. Гуртового. Изд.: «Высшая школа», М., 1978.- 406 с.
6.Гайворонский И.В. Нормальная анатомия человека: учебн. в 2-х т. /И.В.Гайворокский – 3-е изд., исправл. – Спб.:Спецлит, 2003, т.1 – 2003. – 560с, т.2 – 2003. – 424с.
7.Гистология (введение в патологию). Учебник для студентов высших мед. учебных заведений./Под ред. Э.Г.Улумбекова, Ю.А. Челышева. Изд.:«ГЭОТАР», М.,1997.- 947с.
8.Зуссман, М. Биология развития./ Под ред. С.Г. Васецкого. Пер. с анг. Изд.: «Мир». М. 1977-301с.
9.Левина С.Е. Очерки развития пола в раннем онтогенезе высших позвоночных. Изд.: «Наука». М., 1974.-239с.
10.Лейбсон Л.Г. Основные черты структурной и функциональной эволюции эндокринной системы позвоночных. Журн. эвол. биохимии и физиологии, 1967, т.3., №6, с. 532 – 544.
11.Лукин Е.И. Зоология: учебник для студентов зооинженерных и зооветеринарных вузов и факультетов. – 2-е изд., перераб. и дополн. – Изд.:«Высшая школа», 1981, М.- 340 с.
12.Наумов С.П. Зоология позвоночных. Изд.:«Просвещение», М., 1982.-464 с.
13.Талызин Ф.Ф., Улисова Т.Н. Материалы к сравнительной анатомии систем органов позвоночных. Учебное пособие для студентов. М., 1974.-71 с.
14. Физиология человека и животных (общая и эволюционно-экологическая), В 2-х частях. Под ред. Когана А.Б. Изд.: «Высшая школа». М. 1984, I-я часть - 360 с., II часть – 288 с.
15.Шмальгаузен И.И. Основы сравнительной анатомии. Гос. издательство биол. и медицинской литературы. М., 1935.-924 с.
К некоторым вопросам эволюции
Издание 2-е, дополненное
Составители : Ходжаян А.Б., Краснова Л.А., Федоренко Н.Н.
Сдано в набор 05.07.09. Подписано в печать 05.07.09.-Формат 60х99.
Бумага типог. № 1. Печать офсетная. Гарнитура офсетная. Усл. печ. л. 2,0.
Уч.-изд. л. 1,2. Заказ 2087.Тираж 100.
Ставропольская государственная медицинская академия.
355017, г. Ставрополь, ул. Мира, 310.
«Этап 1. Образование центральной нервной системы в виде трубки впервые появляется у животных представителей типа хордовых. У низших хордовых, например у ланцетника, нервная трубка сохраняется в течение всей жизни, у высших хордовых - позвоночных животных - в эмбриональной стадии на спинной стороне зародыша закладывается нервная пластинка, которая погружается под кожу и сворачивается в трубку.
Этап 2. У позвоночных животных нервная трубка подразделяется на головной и спинной мозг. В эмбриональной стадии развития нервная трубка образует в передней части три вздутия - три мозговых пузыря, из которых развиваются отделы мозга: передний пузырь даёт передний и промежуточный мозг, средний пузырь превращается в средний мозг, задний пузырь образует мозжечок и продолговатый мозг. Эти пять отделов мозга характерны для всех позвоночных животных.
Этап 3. Для низших позвоночных - рыб и земноводных - характерно преобладание среднего мозга над остальными отделами. Только у хрящевых акуловых рыб в связи с быстрым движением развит мозжечок, а сильно развитое обоняние привело к увеличению переднего мозга, который становится центром переработки обонятельных сигналов.
Этап 4. У земноводных несколько увеличивается передний мозг и в крыше полушарий образуется тонкий слой нервных клеток - первичный мозговой свод (архипаллиум), древняя кора. Помимо архипаллиума, у земноводных усиливаются связи переднего и среднего мозга.
Этап 5. У рептилий значительно увеличивается передний мозг за счёт скоплений нервных клеток - полосатых тел - на дне переднего мозга. Большую часть крыши полушарий занимает древняя кора. Впервые у рептилий появляется зачаток новой коры - неопаллиум. Полушария переднего мозга наползают на другие отделы, вследствие чего образуется изгиб в области промежуточного мозга. Начиная с древних рептилий, полушария головного мозга становятся самым большим отделом головного мозга.
В строении головного мозга птиц и пресмыкающихся много общего. На крыше головного мозга - первичная кора, хорошо развит средний мозг. Однако у птиц по сравнению с рептилиями возрастают общая масса мозга и относительные размеры переднего мозга. Крупные зрительные доли среднего мозга свидетельствуют об усилении роли зрения в поведении птиц. Мозжечок крупный и имеет складчатое строение.
Значительная часть обоих полушарий переднего мозга у птиц, как и у рептилий, образуют полосатые тела - разрастания дна переднего мозга.
Этап 6. У млекопитающих передний мозг достигает наибольшей величины и сложности. Большую часть мозгового вещества составляет новая кора - вторичный мозговой свод, или неопаллиум. Он состоит из нервных клеток и волокон, расположенных в несколько слоёв. Новая кора больших полушарий служит центром высшей нервной деятельности.
Промежуточный и средний отделы мозга у млекопитающих невелики. Разрастающиеся полушария переднего мозга накрывают их и подминают под себя. У приматов полушария переднего мозга покрывают собой и мозжечок, а у человека и продолговатый мозг. У некоторых млекопитающих мозг гладкий, без борозд и извилин, но у большинства млекопитающих в коре мозга имеются борозды и извилины, которые образуются в процессе роста коры. Наибольшие образования борозд у китообразных, наименьшие - у насекомоядных и летучих мышей.
Этап 7. Появление борозд и извилин происходит вследствие роста мозга при ограниченных размерах черепа. Мозг как бы впечатывается в костные стенки черепа, утесняются оболочки мозга. Дальнейший рост коры приводит к появлению складчатости в виде борозд и извилин. В коре мозга всех млекопитающих находятся ядерные зоны анализаторов, т.е. поля первичного коркового анализа».
Дерягина М.А., Эволюционная антропология: биологические и культурные аспекты, М., Изд-во «УРАО», 2003 г., с. 148-150.
В эволюции нервная система претерпела несколько этапов развития, которые стали поворотными пунктами в качественной организации её деятельности. Эти этапы отличаются по количеству и видам нейрональных образований, синапсов, признакам их функциональной специализации, по образованию группировок нейронов, связанных между собой общностью функций. Выделяют три основных этапа структурной организации нервной системы: диффузный, узловой, трубчатый.
Диффузная нервная система наиболее древняя, имеется у кишечнополостных (гидра) животных. Такая нервная система характеризуется множественностью связей соседних элементов, что позволяет возбуждению свободно распространяться по нервной сети во все стороны.
Этот тип нервной системы обеспечивает широкую взаимозаменяемость и тем самым большую надёжность функционирования, однако эти реакции имеют неточный, расплывчатый характер.
Узловой тип нервной системы типичен для червей, моллюсков, ракообразных.
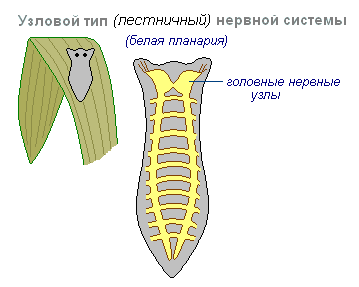
Он характерен тем, что связи нервных клеток организованы определённым образом, возбуждение проходит по жёстко определённым путям. Такая организация нервной системы оказывается более ранимой. Повреждение одного узла вызывает нарушение функций всего организма в целом, но она по своим качествам быстрее и точнее.
Трубчатая нервная система характерна для хордовых, она включает в себя черты диффузного и узлового типов. Нервная система высших животных взяла всё лучшее: высокую надёжность диффузного типа, точность, локальность быстроту организации реакций узлового типа.
Ведущая роль нервной системы
На первом этапе развития мира живых существ взаимодействие между простейшими организмами осуществлялось через водную среду первобытного океана, в которую поступали химические вещества, выделяемые ими. Первой древнейшей формой взаимодействия между клетками многоклеточных организм является химическое взаимодействие посредством продуктов обмена веществ, поступающих в жидкости организма. Такими продуктами обмена веществ, или метаболитами, являются продукты распада белков, углекислота и др. это — гуморальная передача влияний, гуморальный механизм корреляции, или связи между органами.
Гуморальная связь характеризуется следующими особенностями:
- отсутствием точного адреса, по которому направляется химическое вещество, поступающее в кровь или другие жидкости тела;
- химическое вещество распространяется медленно;
- химическое вещество действует в ничтожных количествах и обычно быстро разрушается или выводится из организма.
Гуморальные связи являются общими и для мира животных, и для мира растений. На определённой ступени развития мира животных в связи с появлением нервной системы образуется новая, нервная форма связей и регуляций, которая качественно отличает мир животных от мира растений. Чем выше по своему развитию организм животного, тем большую роль играет взаимодействие органов через нервную систему, которое обозначается как рефлекторное. У высших живых организмов нервная система регулирует гуморальные связи. В отличие от гуморальной связи нервная связь имеет точную направленность к определённому органу и даже группе клеток; связь осуществляется в сотни раз с большей скоростью, чем скорость распространения химических веществ. Переход от гуморальной связи к нервной сопровождался не уничтожением гуморальной связи между клетками тела, а подчинением нервным связям и возникновению нервно-гуморальным связям.
На следующем этапе развития живых существ появляются специальные органы — железы, в которых вырабатываются гормоны, образующиеся из поступающих в организм пищевых веществ. Основная функция нервной системы заключается как в регуляции деятельности отдельных органов между собой, так и во взаимодействии организма как единого целого с окружающей его внешней средой. Любое воздействие внешней среды на организм оказывается, прежде всего, на рецепторы (органы чувств) и осуществляется через посредство изменений, вызываемых внешней средой и нервной системой. По мере развития нервной системы высший её отдел — большие полушария головного мозга — становится «распорядителем и распределителем всей деятельности организма».
Строение нервной системы
Нервная система образована нервной тканью, которая состоит из огромного количества нейронов — нервная клетка с отростками.

Нервная система условно подразделяется на центральную и периферическую.

Центральная нервная система включает головной и спинной мозг, а периферическая нервная система - нервы, отходящие от них.

Головной и спинной мозг представляют собой совокупность нейронов. На поперечном разрезе мозга различают белое и серое вещество. Серое вещество состоит из нервных клеток, а белое - из нервных волокон, являющихся отростками нервных клеток. В различных отделах центральной нервной системы расположение белого и серого вещества неодинаково. В спинном мозге серое вещество находится внутри, а белое — снаружи, в головном же (большие полушария, мозжечок), наоборот — серое вещество — снаружи, белое — внутри. В различных отделах головного мозга имеются отдельные скопления нервных клеток (серого вещества), расположенные внутри белого вещества, - ядра . Скопления нервных клеток находятся и за пределами центральной нервной системы. Они называются узлами и относятся к периферической нервной системе.
Рефлекторная деятельность нервной системы
Основной формой деятельности нервной системы является рефлекс. Рефлекс - реакция организма на изменение внутренней или внешней среды, осуществляемая при участии центральной нервной системы в ответ на раздражение рецепторов.

При всяком раздражении возбуждение с рецепторов передаётся по центростремительным нервным волокнам в центральную нервную систему, откуда через вставочный нейрон по центробежным волокнам оно идёт на периферию к тому или иному органу, деятельность которого изменяется. Весь этот путь через центральную нервную систему к рабочему органу, называется рефлекторной дугой образован обычно тремя нейронами: чувствительным, вставочным и двигательным. Рефлекс — сложный акт, в осуществлении которого принимает участие значительно большее количество нейронов. Возбуждение, попадая в центральную нервную систему, распространяется на многие отделы спинного мозга и доходит до головного. В результате взаимодействия многих нейронов осуществляется ответная реакция организма на раздражение.
Спинной мозг
Спинной мозг - тяж длиной около 45 см, диаметром 1 см, находится в канале позвоночника, покрыт тремя мозговыми оболочками: твёрдой, паутинной и мягкой (сосудистой).
Спинной мозг находится в позвоночном канале и представляет собой тяж, который вверху переходит в продолговатый мозг, а внизу заканчивается на уровне второго поясничного позвонка. Спинной мозг состоит из серого вещества, содержащего нервные клетки, и белого, состоящего из нервных волокон. Серое вещество расположено внутри спинного мозга и окружено со всех сторон белым веществом.
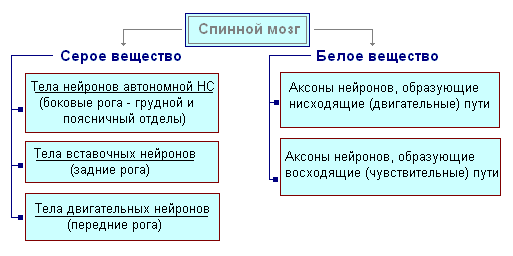
На поперечном разрезе серое вещество напоминает букву Н. В нём различают передние и задние рога, а также соединяющую перекладину, в центре которой находится узкий канал спинного мозга, содержащий спинномозговую жидкость. В грудном отделе выделяют боковые рога. В них заложены тела нейронов, иннервирующих внутренние органы. Белое вещество спинного мозга образовано нервными отростками. Короткие отростки соединяют участки спинного мозга, а длинные составляют проводниковый аппарат двусторонних связей с головным мозгом.
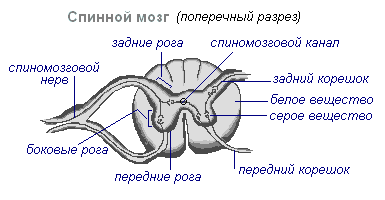
Спинной мозг имеет два утолщения - шейное и поясничное, от которых отходят нервы к верхним и нижним конечностям. От спинного мозга отходит 31 пара спинномозговых нервов. Каждый нерв начинается от спинного мозга двумя корешками — передним и задним. Задние корешки — чувствительные состоят из отростков центростремительных нейронов. Их тела расположены в спинномозговых узлах. Передние корешки — двигательные — являются отростками центробежных нейронов расположенных в сером веществе спинного мозга. В результате слияния переднего и заднего корешка образуется смешанный спинномозговой нерв. В спинном мозге сосредоточены центры, регулирующие наиболее простые рефлекторные акты. Основные функции спинного мозга - рефлекторная деятельность и проведение возбуждения.
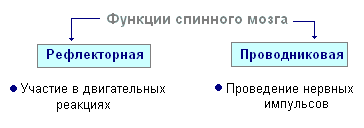
В спинном мозге человека заложены рефлекторные центры мышц верхних и нижних конечностей, потоотделения и мочеиспускания. Функции проведения возбуждения заключается в том, что через спинной мозг проходят импульсы от головного мозга ко всем областям тела и обратно. По восходящим проводящим путям в головной мозг передаются центростемительные импульсы от органов (кожа, мышцы). По нисходящим путям центробежные импульсы передаются от головного мозга в спинной, затем на периферию, к органам. При повреждении проводящих путей наблюдается потеря чувствительности в различных участках тела, нарушение произвольных сокращений мышц и способности к движению.
Эволюция головного мозга позвоночных
Образование центральной нервной системы в виде нервной трубки впервые появляется у хордовых. У низших хордовых нервная трубка сохраняется в течение всей жизни, у высших - позвоночных - в стадии эмбриона на спинной стороне закладывается нервная пластинка, которая погружается под кожу и сворачивается в трубку. В эмбриональной стадии развития нервная трубка образует в передней части три вздутия — три мозговых пузыря, из которых развиваются отделы мозга: передний пузырь дает передний и промежуточный мозг, средний пузырь превращается в средний мозг, задний пузырь образует мозжечок и продолговатый мозг . Эти пять отделов мозга характерны для всех позвоночных животных.
Для низших позвоночных - рыб и земноводных - характерно преобладание среднего мозга над остальными отделами. У земноводных несколько увеличивается передний мозг и в крыше полушарий образуется тонкий слой нервных клеток - первичный мозговой свод, древняя кора. У рептилий значительно увеличивается передний мозг за счет скоплений нервных клеток. Большую часть крыши полушарий занимает древняя кора. Впервые у рептилий появляется зачаток новой коры. Полушария переднего мозга наползают на другие отделы, вследствие чего образуется изгиб в области промежуточного мозга. Начиная с древних рептилий, полушария головного мозга становятся самым большим отделом головного мозга.
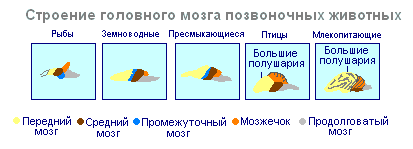
В строении головного мозгаптиц и пресмыкающихся много общего. На крыше головного мозга - первичная кора, хорошо развит средний мозг. Однако у птиц по сравнению с рептилиями возрастают общая масса мозга и относительные размеры переднего мозга. Мозжечок крупный и имеет складчатое строение. У млекопитающих передний мозг достигает наибольшей величины и сложности. Большую часть мозгового вещества составляет новая кора, которая служит центром высшей нервной деятельности. Промежуточный и средний отделы мозга у млекопитающих невелики. Разрастающиеся полушария переднего мозга накрывают их и подминают под себя. У некоторых млекопитающих мозг гладкий, без борозд и извилин, но у большинства млекопитающих в коре мозга имеются борозды и извилины. Появление борозд и извилин происходит вследствие роста мозга при ограниченных размерах черепа. Дальнейший рост коры приводит к появлению складчатости в виде борозд и извилин.
Головной мозг
Если спинной мозг у всех позвоночных животных развит более или менее одинаково, то головной мозг существенно отличатся размерами и сложностью строения у разных животных. Особенно резкие изменения в ходе эволюции претерпевает передний мозг. У низших позвоночных передний мозг развит слабо. У рыб он представлен обонятельными долями и ядрами серого вещества в толще мозга. Интенсивное развитие переднего мозга связано с выходом животных на сушу. Он дифференцируется на промежуточный мозг и на два симметричных полушария, которые называются конечным мозгом . Серое вещество на поверхности переднего мозга (кора) впервые появляется у пресмыкающихся, развиваясь далее у птиц и особенно у млекопитающих. Действительно большими полушариями переднего мозга становятся только у птиц и млекопитающих. У последних они покрывают почти все другие отделы головного мозга.
Головной мозг расположен в полости черепа. В него входят ствол и конечный мозг (кора больших полушарий).

Ствол мозга состоит из продолговатого мозга, варолиева моста, среднего и промежуточного мозга.
Продолговатый мозг является непосредственным продолжением спинного мозга и расширяясь, переходит в задний мозг. Он в основном сохраняет форму и строение спинного мозга. В толще продолговатого мозга расположены скопления серого вещества — ядра черепно-мозговых нервов. В состав заднего моста входят мозжечок и варолиев мост . Мозжечок расположен над продолговатым мозгом и имеет сложное строение. На поверхности полушарий мозжечка серое вещество образует кору, а внутри мозжечка - его ядра. Как и спинной продолговатый мозг выполняет две функции: рефлекторную и проводниковую. Однако рефлексы продолговатого мозга более сложные. Это выражается в важном значении в регуляции сердечной деятельности, состоянии сосудов, дыхания, потоотделения. В продолговатом мозге расположены центры всех этих функций. Здесь же находятся центры жевания, сосания, глотания, отделения слюны и желудочного сока. Несмотря на малую величину (2,5–3 см), продолговатый мозг представляет собой жизненно важный отдел ЦНС. Повреждение его может стать причиной смерти вследствие прекращения дыхания и деятельности сердца. Проводниковая функция продолговатого мозга и варолиева моста заключается в передаче импульсов из спинного мозга в головной и обратно.
В среднем мозге расположены первичные (подкорковые) центры зрения и слуха, которые осуществляют рефлекторные ориентировочные реакции на световые и звуковые раздражения. Эти реакции выражаются в различных движениях туловища, головы и глаз в сторону раздражителей. Средний мозг состоит из ножек мозга и четверохолмия. Средний мозг регулирует и распределяет тонус (напряжение) скелетных мышц.
Промежуточный мозг состоит из двух отделов - таламус и гипоталамус , каждый из которых состоит из большого числа ядер зрительных бугров и подбугровой области. Через зрительные бугры центростремительные импульсы передаются к коре больших полушарий от всех рецепторов тела. Ни один центростремительный импульс, откуда бы он ни шёл, не может пройти к коре, минуя зрительные бугры. Таким образом, через промежуточный мозг осуществляется связь всех рецепторов с корой больших полушарий. В подбугровой области расположены центры, оказывающие влияние на обмен веществ, терморегуляцию и железы внутренней секреции.
Мозжечок находится позади продолговатого мозга. Он состоит из серого и белого вещества. Однако в отличие от спинного мозга и ствола серое вещество - кора - находится на поверхности мозжечка, а белое вещество расположено внутри, под корой. Мозжечок координирует движения, делает их чёткими и плавными, играет важную роль в сохранении равновесия тела в пространстве, а также оказывает влияние на тонус мышц. При поражении мозжечка у человека наблюдается падение тонуса мышц, расстройство движений и изменение походки, замедляется речь и т.д. Однако через некоторое время движения и мышечный тонус восстанавливаются благодаря тому, что неповреждённые участки центральной нервной системы берут на себя функции мозжечка.
Большие полушария - наиболее крупный и развитый отдел головного мозга. У человека они образуют основную массу головного мозга и по всей своей поверхности покрыты корой. Серое вещество покрывает полушария снаружи и образует кору головного мозга. Кора полушарий человека имеет толщину от 2 до 4 мм и слагается из 6–8 слоёв, образованных 14–16 млрд. клеток, различных по форме, величине и выполняемым функциям. Под корой находится белое вещество. Оно состоит из нервных волокон, связывающих кору с расположенными ниже отделами центральной нервной системы и отдельные доли полушарий между собой.
Кора головного мозга имеет извилины, разделённые бороздами, которые значительно увеличивают её поверхность. Три самые глубокие борозды делят полушария на доли. В каждом полушарии различают четыре доли: лобную, теменную, височную, затылочную . Возбуждение разных рецепторов поступают в соответствующие воспринимающие участки коры, называемые зонами , и отсюда передаются к определённому органу, побуждая его к действию. В коре выделяют следующие зоны. Слуховая зона расположена в височной доле, воспринимает импульсы от слуховых рецепторов.

Зрительная зона лежит в затылочной области. Сюда поступают импульсы от рецепторов глаза.
Обонятельная зона находится на внутренней поверхности височной доли и связана с рецепторами носовой полости.
Чувствительно-двигательная зона расположена в лобной и теменной долях. В этой зоне находятся главные центры движения ног, туловища, рук, шеи, языка и губ. Здесь же лежит и центр речи.

Полушария головного мозга - это высший отдел центральной нервной системы, контролирующий работу всех органов у млекопитающих. Значение больших полушарий у человека заключается ещё и в том, что они представляют собой материальную основу психической деятельности. И.П.Павлов показал, что в основе психической деятельности лежат физиологические процессы, происходящие в коре головного мозга. Мышление связано с деятельностью всей коры головного мозга, а не только с функцией отдельных её областей.
| Отдел головного мозга | Функции | |
| Продолговатый мозг | Проводниковая | Связь спинного и вышележащих отделов головного мозга. |
| Рефлекторная | Регуляция деятельности дыхательной, сердечно-сосудистой, пищеварительной систем:
|
|
| Варолиев мост | Проводниковая | Соединяет полушария мозжечка между собой и с корой больших полушарий головного мозга. |
| Мозжечок | Координационная | Координация произвольных движений и сохранение положения тела в пространстве. Регуляция мышечного тонуса и равновесия |
| Средний мозг | Проводниковая | Ориентировочные рефлексы на зрительные, звуковые раздражители (повороты головы и туловища ). |
| Рефлекторная |
|
|
| Промежуточный мозг | таламус
гипоталамус
|
|
Кора больших полушарий
Поверхность коры больших полушарий у человека составляет около 1500 см 2 , что во много раз превышает внутреннюю поверхность черепа. Такая большая поверхность коры образовалась благодаря развитию большого количества борозд и извилин, в результате чего большая часть коры (около 70%) сосредоточена в бороздах. Самые большие борозды больших полушарий - центральная , которая проходит поперёк обоих полушарий, и височная , отделяющая височную долю от остальных. Кора больших полушарий, несмотря на малую толщину (1,5–3 мм) имеет очень сложное строение. В ней насчитывают шесть основных слоёв, которые отличаются строением, формой и размерами нейронов и связями. В коре находятся центры всех чувствительных (рецепторных) систем, представительства всех органов и частей тела. В связи с этим к коре подходят центростремительные нервные импульсы от всех внутренних органов или частей тела, и она может управлять их работой. Через кору больших полушарий происходит замыкание условных рефлексов, посредством которых организм постоянно, в течение всей жизни очень точно приспосабливается к изменчивым условиям существования, к окружающей среде.
Антропология и концепции биологии Курчанов Николай Анатольевич
Эволюция мозга позвоночных
Эволюция мозга позвоночных
При сравнительно-анатомическом анализе строения мозга позвоночных наибольшее впечатление производит эволюция полушарий большого мозга. Возникнув как выросты переднего мозга, исключительно в связи с обонятельной рецепцией, они становятся основным ассоциативным центром у представителей высокоорганизованных классов (Ромер А., Парсонс Т., 1992). В эволюции большого мозга важнейшим этапом было образование коры , которая появляется уже у представителей класса пресмыкающихся. Рептилии представляют собой тот узловой этап, от которого формируются две независимые филогенетические линии эволюции мозга: стриарная , представленная птицами, и кортикальная , представленная млекопитающими. В очередной раз отметим, что эволюция характеризуется не линейной направленностью, а дивергенцией.
У птиц кора развита слабо, а неокортекс вовсе отсутствует. Эволюция их мозга характеризуется совершенствованием сложной системы стереотипных реакций, т. е. реализацией стратегии ригидности. Но многие птицы проявляют неплохие способности к обучению на основе индивидуального опыта. Резко выделяются в этом плане врановые и попугаи. Однако способности птиц к обучению связаны не с корой. Их контролирует особая область полосатого тела – гиперстриатум , аналогичный по своим функциям неокортексу млекопитающих (Ромер А., Парсонс Т., 1992).
У млекопитающих высшие интегральные функции концентрируются в коре полушарий большого мозга. Основную часть коры мозга млекопитающих составляет неокортекс (новая кора), возникающий уже у высших рептилий. Эволюция мозга млекопитающих – это, по сути дела, эволюция неокортекса. Новая кора млекопитающих не только становится больше других отделов мозга, но и берет на себя функции координатора вновь возникающих центров высшей нервной деятельности. Особенно впечатляющих размеров неокортекс достигает у человека: 99 % всех его нейронов составляют вставочные нейроны, образующие гигантское число связей (Хамори Й., 1985).
Параллелизм в эволюции когнитивных функций у позвоночных с разными типами структурно-функциональной организации мозга показывает, что эволюция мозга в разных группах шла независимыми путями с различной скоростью. Поэтому ряд авторов обращают внимание на то, что нельзя переносить наблюдения взаимосвязи структуры и функции отделов мозга с одной таксономической группы на другую (Hodos W., 1982).
В настоящее время существует несколько гипотез эволюции мозга позвоночных, но единой теории пока нет.
Данный текст является ознакомительным фрагментом. Из книги Исчезнувший мир автора Акимушкин Игорь ИвановичНачало эволюции позвоночных Ланцетник - реликт, живое ископаемое, чудом сохранившееся от давно минувших дней и поныне еще роется в песке на мелководьях теплых морей. Впрочем не совсем так. Некоторые из этих странных создании на дне не живут.«Ланцетники амфиоксиды…
Из книги Основы зоопсихологии автора Фабри Курт ЭрнестовичПостэмбриональное развитие поведения у зрело- и незрелорождающихся позвоночных Детеныши позвоночных рождаются на разных стадиях зрелости. В предыдущей главе мы уже познакомились с примером крайнего незрелорождения - кенгуренком, появляющимся на свет еще в
Из книги Новейшая книга фактов. Том 1 [Астрономия и астрофизика. География и другие науки о Земле. Биология и медицина] автораФормирование общения у низших позвоночных Отмеченные закономерности не имеют конечно, значения всеобщих правил для всех позвоночных, а тем более беспозвоночных. Даже среди птиц и млекопитающих обнаруживаются многочисленные отклонения и исключения, обусловленные
Из книги Новейшая книга фактов. Том 1. Астрономия и астрофизика. География и другие науки о Земле. Биология и медицина автора Кондрашов Анатолий ПавловичРигидность и пластичность в поведении высших позвоночных Как уже указывалось, вопреки еще распространенному мнению инстинктивное поведение не теряет своей значимости в процессе эволюции, так как оно принципиально не может замещаться научением. Подчеркнем еще раз, что
Из книги Мозг, разум и поведение автора Блум Флойд Э Из книги Эволюция [Классические идеи в свете новых открытий] автора Марков Александр ВладимировичУ кого из позвоночных самые большие глаза? Самое крупное глазное яблоко среди всех позвоночных принадлежало ихтиозавру, хотя он был далеко не самым крупным животным. Этот ящер, внешне напоминавший тунца или дельфина длиной до 15 метров, нырял в поисках пищи на глубины до 600
Из книги Удивительная палеонтология [История земли и жизни на ней] автора Еськов Кирилл Юрьевич5. Ритмы мозга Мы без труда замечаем ритмические изменения, происходящие в окружающем нас мире: весна, лето, осень и зима образуют привычный цикл; солнце всходит каждый день, движется по небу и садится; луна прибывает и убывает; в океане приливы чередуются с отливами.
Из книги Происхождение мозга автора Савельев Сергей ВячеславовичСекретный код позвоночных выдал ланцетник Ланцетник (Branchiostoma) - хрестоматийный пример живого ископаемого, т. е. мало изменившегося потомка очень древнего предка. Ланцетника называют живой схемой хордового животного. Это маленькое червеобразное существо большую часть
Из книги Расы. Народы. Интеллект [Кто умнее] автора Линн РичардГЛАВА 10 Поздний палеозой - ранний мезозой: эволюция наземных позвоночных (1). Анамнии и амниоты. Две линии амниот - тероморфная и завроморфная Мы расстались с самыми первыми тетраподами (девонской ихтиостегой и родственными ей формами), когда те… Чуть было не сказал:
Из книги Антропология и концепции биологии автораГЛАВА 11 Поздний мезозой: эволюция наземных позвоночных (2). Завроморфный мир. Маммализация териодонтов. Динозавры и их вымирание Итак, мы знаем, что с самого момента появления истинно наземных позвоночных - амниот - произошло их разделение на две эволюционные ветви:
Из книги Поведение: эволюционный подход автора Курчанов Николай Анатольевич§ 12. Нервная система позвоночных Нервная система позвоночных построена на принципах вероятностного развития, дублирования, избыточности и индивидуальной изменчивости. Это не означает, что в мозге позвоночных нет места генетической детерминации развития или
Из книги автора§ 27. Мозг первичноводных позвоночных Рассмотрим основные принципы строения универсальной конструкции нервной системы первичноводных позвоночных (рис. II-17; II-18). Её морфологической осью является центральная нервная система, которая расположена над позвоночником (рис.
Из книги автора§ 29. Формирование мозга позвоночных Теперь попробуем предположить те события, которые могли привести к развитию описанной выше конструкции головного мозга древних первичноводных позвоночных. Жизнь в мелководной билатерали должна была иметь существенные преимущества
Из книги автора2. Размер мозга Размер мозга бушменов был оценен в l250 см 3Дреннаном (Drennan; 1937) и немного выше – в l270 см 3Смитом и Билсом (Smith, Beals; 1990). Массив данных Смита и Билса включал также африканцев-негроидов, размер мозга которых составил l282 см 3, то есть немного больше,
Из книги автора10.1. Эволюция мозга Как сказал один из творцов современной молекулярной биологии, лауреат Нобелевской премии Ф. Крик: «Нет области науки более жизненно важной для человека, чем исследования его собственного мозга» (Крик Ф.,
Из книги автора8.3. Основные тенденции эволюции нервной системы позвоночных Позвоночные обычно рассматриваются как подтип Vertebrata в типе хордовых (Chordata). Они представлены бесчелюстными, рыбами, амфибиями, рептилиями, птицами и млекопитающими. В систематике позвоночных много спорных
Нервная система хордовых животных, как и у всех многоклеточных, развивается из эктодермы. Она возникла за счет погружения чувствительных клеток, первоначально лежавших на поверхности тела, под его покровы. Это доказывается и сравнительно-анатомическими, и эмбриологическими данными.
Действительно, у наиболее примитивного представителя хордовых - ланцетника -центральная нервная система, состоящая из нервной трубки, сохранила функции органа чувств: среди клеток, лежащих внутри нее, имеются отдельные светочувствительные образования - глазки Гессе.Кроме того, основные дистантные органы чувств -зрения, обоняния и слуха -образуются у всех позвоночных первоначально как выпячивания передней части нервной трубки.
В эмбриогенезе нервная система формируется вначале всегда в виде полосы утолщенной эктодермы на спинной стороне зародыша, которая впячивается под покровы и замыкается в трубку с полостью внутри - невроцелем. У ланцетника это замыкание еще не полное, поэтому нервная трубка выглядит как желобок (рис. 14.38).Передний конец ее расширен. Он гомологичен головному мозгу позвоночных. Большинство клеток нервной трубки ланцетника не являются нервными, они выполняют опорные или рецепторные функции.
Рис 14.38.Нервная трубка ланцетника: 1 -невроцель, 2 - глазки Гессе
У всех позвоночных центральная нервная система является производной нервной трубки, передний конец которой становится головным мозгом, а задний -спинным. Образование головного мозга называюткефализацией. Она связана с усилением двигательной активности позвоночных и необходимостью постоянного анализа раздражении, приходящих из внешней среды, в первую очередь с переднего конца тела. Этот процесс сопровождается также дифференциацией органов чувств, особенно дистантных -обоняния, зрения и слуха. Совместная эволюция органов чувств и головного мозга приводит к возникновению динамических координации между обонятельными рецепторами и передним мозгом, зрительными -и средним, слуховыми -и задним. Видимо, поэтому головной мозг всех современных позвоночных животных в эмбриогенезе закладывается вначале из трех мозговых пузырей -переднего, среднего и заднего -и только позже дифференцируется на пять отделов. Вероятно, предки позвоночных имели более простой головной мозг, развивающийся на основе трех мозговых пузырей.
Головной мозгсовременных взрослых позвоночных всегда состоит из пяти отделов: переднего, промежуточного, среднего, заднего и продолговатого. Внутри головного и спинного мозга расположена общая полость, соответствующая невроцелю. В спинном мозге этоспинномозговой канал,а в головном -желудочки мозга.Ткань мозга состоит изсерого вещества (скопления нервных клеток) ибелого (отростков нервных клеток).
Во всех отделах головного мозга различают мантию, располагающуюся над желудочками, и основание, лежащее под ними. В прогрессивной эволюции головного мозга проявляется постепенное усиление роли его передних отделов и мантии по сравнению с задними и основанием.
У рыб головной мозг в целом невелик. Слабо развит его передний отдел. Передний мозг не разделен на полушария. Крыша его тонкая, состоит только из эпителиальных клеток и не содержит нервной ткани. Основание переднего мозга включает полосатые тела, от него отходят обонятельные доли. Функционально передний мозг является высшим обонятельным центром.
В промежуточном мозге, с которым связаны эпифиз и гипофиз, расположен гипоталамус, являющийся центральным органом эндокринной системы. Средний мозг рыб наиболее развит. Он состоит из двух полушарий и служит высшим зрительным центром. Кроме того, он представляет собой высший интегрирующий отдел головного мозга. Задний мозг содержит мозжечок, осуществляющий регуляцию координации движений. Он развит очень хорошо в связи с перемещением рыб в трехмерном пространстве. Продолговатый мозг обеспечивает связь высших отделов головного мозга со спинным и содержит центры дыхания и кровообращения. Головной мозг такого типа, в котором высшим центром интеграции функций является средний мозг, называют ихтиопсидным.
У земноводных головной мозг также ихтиопсидный. Однако передний мозг их имеет большие размеры и разделен на полушария. Крыша его состоит из нервных клеток, отростки которых располагаются на поверхности. Как и у рыб, больших размеров достигает средний мозг, также представляющий собой высший интегрирующий центр и центр зрения. Мозжечок несколько редуцирован в связи с примитивным характером движений.
Условия наземного существования пресмыкающихся требуют более сложной морфофункциональной организации мозга. Передний мозг -наиболее крупный отдел по сравнению с остальными. В нем особенно развиты полосатые тела. К ним переходят функции высшего интегративного центра. На поверхности крыши впервые появляются островки коры очень примитивного строения, ее называютдревней - archicortex. Средний мозг теряет значение ведущего отдела, и относительные размеры его сокращаются. Мозжечок сильно развит благодаря сложности и многообразию движений пресмыкающихся. Головной мозг такого типа, в котором ведущий отдел представлен полосатыми телами переднего мозга, называютзауропсидным.
У млекопитающих -маммалийный тип мозга. Для него характерно сильное развитие переднего мозга за счет коры, которая развивается на основе небольшого островка коры пресмыкающихся и становится интегрирующим центром мозга. В ней располагаются высшие центры зрительного, слухового, осязательного, двигательного анализаторов, а также центры высшей нервной деятельности. Кора имеет очень сложное строение и называетсяновой корой - neocortex. В ней располагаются не только тела нейронов, но и ассоциативные волокна, соединяющие разные ее участки. Характерным является также наличие комиссуры между обоими полушариями, в которой располагаются волокна, связывающие их воедино. Промежуточный мозг, как и у других классов, включает гипоталамус, гипофиз и эпифиз. В среднем мозге располагается четверохолмие в виде четырех бугров. Два передних связаны со зрительным анализатором, два задних-со слуховым. Очень хорошо развит мозжечок (рис. 14.39).

Рис. 14.39.Эволюция головного мозга позвоночных. А-рыба;Б- земноводное;В- пресмыкающееся;Г- млекопитающее:
1 -обонятельные доли, 2- передний мозг, 3- средний мозг, 4- мозжечок, 5- продолговатый мозг, 6- промежуточный мозг
По мере усиления функций передних отделов головного мозга в филогенезе спинного мозга наблюдается его продольная дифференцировка с образованием утолщений в области отхождения крупных нервов к конечностям и редукция его заднего конца.
Так, у рыб спинной мозг равномерно тянется вдоль всего тела. Начиная от земноводных происходит его укорочение сзади. У млекопитающих на заднем конце спинного мозга остается рудимент в виде конечной нити -filumterminale. Нервы, идущие к заднему концу тела, проходят по позвоночному каналу самостоятельно, образуя так называемый конский хвост -caudaequina.
Основные этапы эволюции центральной нервной системы отражаются и в онтогенезе человека. На стадии нейруляции закладывается нервная пластинка, превращающаяся в желобок и затем в трубку. Передний конец трубки образует сначала три мозговых пузыря (рис. 14.40): передний (I ), средний (II )и задний (III ). Вслед за этим передний пузырь подразделяется на два, дифференцирующихся на передний (7) и промежуточный (2 )мозг -telencephalon,diencephalon. Средний мозговой пузырь развивается в средний (3)мозг -mesencephalon,aзадний -в задний (4 )мозг -metencephalon- и продолговатый (5) мозг -medullaoblongata.

Рис. 14.40.Мозговые пузыри в эмбриогенезе мозга человека.А - стадия трех мозговых пузырей;Б - стадия пяти мозговых пузырей (остальные пояснения см. в тексте)
Задний конец спинного мозга редуцируется, превращаясь в терминальную нить. Позже скорости роста спинного мозга и позвоночника оказываются разными, и к моменту рождения конец спинного мозга оказывается на уровне третьего, а у взрослого человека -уже на уровне первого поясничного позвонка.
Головной мозг рано начинает развиваться по пути, характерному для млекопитающих и человека. Первично почти прямая нервная трубка резко изгибается в области будущего продолговатого и среднего мозга. На этом фоне большие полушария переднего мозга растут с особенно большой скоростью. В результате головной мозг оказывается расположенным над лицевым черепом. Дифференцировка коры приводит к развитию извилин, борозд и формированию высших сенсорных и двигательных центров, в том числе центров письменной и устной речи и др., характерных только для человека (см. § 15.3).
Центральная нервная система столь важна для интеграции индивидуального развития человека, что большинство ее врожденных пороков несовместимы с жизнью. Среди пороков спинного мозга, онтогенетические механизмы которых известны, отметим рахисхиз,нли платиневрию,- отсутствие замыкания нервной трубки (рис. 14.41).Эта аномалия связана с нарушением клеточных перемещений и адгезии в зоне формирования нервной трубки в процессе нейруляции. Аномалия переднего мозга -прозэнцефалия - выражается в нарушении морфо-генеза мозга, при котором полушария оказываются неразделенными, а кора -недоразвита. Этот порок формируется на 4-й неделе эмбриогенеза, в момент закладки переднего мозга. Как и предыдущий, он несовместим с жизнью. Часто встречается у мертворожденных при различных хромосомных и генных синдромах.
Нарушения дифференцировки коры -агирия (отсутствие извилин) иолигогирия спахигирией (малое количество утолщенных извилин) - сопровождаются упрощением гистологического строения коры. У детей с такими пороками выявляются грубая олигофрения и нарушение многих рефлексов. Большинство детей умирают в течение первого года жизни.

Рис. 14.41.Рахисхиз (платиневрия)